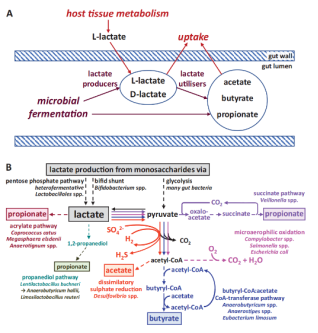
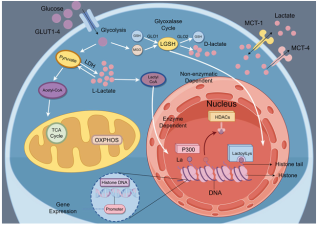
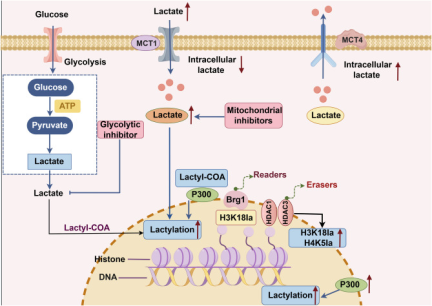
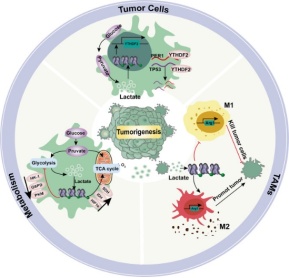
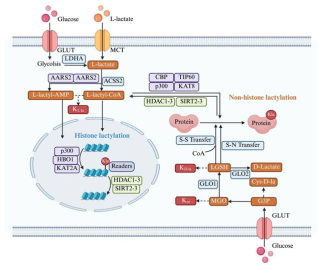
Lactate has long been regarded as a metabolic end product of glycolysis. However, recent studies have revealed its multiple functions in energy supply, signal transduction, and epigenetic regulation. This article systematically reviews the production, transport, and metabolic mechanisms of lactate, with an emphasis on its biological roles in immune regulation, inflammation control, intestinal homeostasis maintenance, and tumor microenvironment remodeling. In addition, it provides an in-depth discussion of the "writers-erasers-readers" regulatory network of lactylation. Lactylation is a lactate-driven post-translational modification that occurs on both histone and non-histone proteins. Histone lactylation primarily regulates gene transcription by altering chromatin conformation and plays key roles in macrophage polarization, inflammation resolution, and tumor progression. Non-histone lactylation extensively modifies various proteins, including metabolic enzymes and transcription factors, and participates in signal transduction, immune escape, and DNA repair. The two types of lactylation exhibit significant differences in modification sites, reaction mechanisms, and functional effects, together forming an important bridge between metabolism and epigenetic regulation. Current studies suggest that targeting lactate metabolism or lactylation-related regulatory pathways holds therapeutic potential in tumors, inflammatory diseases, and metabolic disorders. Future efforts are needed to further elucidate the dynamic regulatory networks of lactylation under pathological conditions and to facilitate its clinical translation.
| Published in | Science Research (Volume 14, Issue 3) |
| DOI | 10.11648/j.sr.20261403.11 |
| Page(s) | 62-70 |
| Creative Commons |
This is an Open Access article, distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution and reproduction in any medium or format, provided the original work is properly cited. |
| Copyright |
Copyright © The Author(s), 2026. Published by Science Publishing Group |
Lactate, Lactylation, Histone, Non-histone
| [1] | Zhang D, Tang Z, Huang H, et al. Metabolic regulation of gene expression by histone lactylation [J]. Nature, 2019, 574(7779): 575-580. |
| [2] | Ren H, Tang Y, Zhang D. The emerging role of protein l-lactylation in metabolic regulation and cell signalling [J]. Nature Metabolism, 2025, 7(4): 647-664. |
| [3] | Huang B, Jin M, Cui G, et al. GCN5–ERK lactylation–phosphorylation loop amplifies lactate-driven cancer progression [J]. Nat Chem Biol, 2026, 22(4): 634-648. |
| [4] | Zong Z, Xie F, Wang S, et al. Alanyl-tRNA synthetase, AARS1, is a lactate sensor and lactyltransferase that lactylates p53 and contributes to tumorigenesis [J]. Cell, 2024, 187(10): 2375-2392. |
| [5] | Peng X, Du J. Histone and non-histone lactylation: molecular mechanisms, biological functions, diseases, and therapeutic targets [J]. Molecular Biomedicine, 2025, 6(38): 1-41. |
| [6] | Chen J, Huang Z, Chen Y, et al. Lactate and lactylation in cancer [J]. Signal Transduction and Targeted Therapy, 2025, 38(3): 1316-1341. |
| [7] | Zhang S, Wang J, Xu Z, et al. Lactate: elucidating its indispensable role in human health [J]. Molecular Cancer, 2026, 25(2): 1-41. |
| [8] | Zong Z, Ren J, Yang B. et al. Emerging roles of lysine lactyltransferases and lactylation [J]. Nat Cell Biol, 2025, 27(4): 563-574. |
| [9] | Cai Q, Deng W, Zou Y, et al. Histone lactylation as a driver of metabolic reprogramming and immune evasion [J]. Med. Rev. 2025, 5(3): 256-259. |
| [10] | Vander Heiden M G, Cantley L C, Thompson C B. Understanding the Warburg effect: the metabolic requirements of cell proliferation [J]. Science, 2009, 324(5930): 1029-1033. |
| [11] | Brown T P, Ganapathy V. Lactate/GPR81 signaling and proton motive force in cancer: Role in angiogenesis, immune escape, nutrition, and Warburg phenomenon [J]. Pharmacology & therapeutics, 2020, 206: 107451. |
| [12] | Liu C, Wu J, Zhu J, et al. Lactate inhibits lipolysis in fat cells through activation of an orphan g-protein-coupled receptor, gpr81 [J]. Journal of Biological Chemistry, 2009, 284(5): 2811-2822. |
| [13] | Bangar S P, Suri S, Trif M, et al. Organic acids production from lactic acid bacteria: A preservation approach [J]. Food Bioscience, 2022, 2(46): 959-974. |
| [14] | Duncan S H, Louis P, Flint H J. Lactate-utilizing bacteria, isolated from human feces, that produce butyrate as a major fermentation product [J]. Appl Environ Microbiol, 2004, 70(10): 5810-5817. |
| [15] | Duncan S H, Reichardt N, Young P, et al. Phylogenetic distribution of three pathways for propionate production within the human gut microbiota [J]. The ISME journal emultidisciplinary journal of microbial ecology, 2014, 8(6): 1323–1335. |
| [16] | Louis P, Flint H J. Formation of propionate and butyrate by the human colonic microbiota [J]. Environmental Microbiology, 2017, 19(1): 29-41. |
| [17] | Louis P, Hold G L, Flint H J. The gut microbiota, bacterial metabolites and colorectal cancer [J]. Nature Reviews Microbiology, 2014, 12(10): 661-672. |
| [18] | Louis P, Duncan S H, Sheridan P O, et al. Microbial lactate utilisation and the stability of the gut microbiome [J]. Gut Microbiome, 2022, 3(e3): 1-16. |
| [19] | Felmlee M A, Jones R S, Rodriguezcruz V, et al. Monocarboxylate Transporters (SLC16): Function, regulation, and role in health and disease [J]. Pharmacological Reviews, 2020, 72(2): 466-485. |
| [20] | Llibre A, Kucuk S, Gope A, Certo M, Mauro C. Lactate: A key regulator of the immune response [J]. Immunity. 2025, 58(3): 535-554. |
| [21] | Zhang J, Wu D, Zeng F, et al. Lactate metabolic reprogramming and histone lactylation modification in sepsis [J]. International Journal of Biological Sciences 2025; 21(11) 5034-5055. |
| [22] | Li X, Yang Y, Zhang B, et al. Lactate metabolism in human health and disease [J]. Signal Transduction and Targeted Therapy, 2022, 7(305): 1-22. |
| [23] | Cai X, Ng C P, Jones O, et al. Lactate activates the mitochondrial electron transport chain independently of its metabolism [J]. Molecular Cell, 2023, 83(21): 3904-3920. |
| [24] | Dittoe D K, Ricke S C, Kiess A S. Organic acids and potential for modifying the avian gastrointestinal tract and reducing pathogens and disease [J]. Frontiers in Veterinary Science, 2018, 5(216): 1-12. |
| [25] | Keersmaecker S C J D, Verhoeven T L A, Desair J, et al. Strong antimicrobial activity of Lactobacillus rhamnosus GG against Salmonella typhimurium is due to accumulation of lactic acid [J]. Fems Microbiology Letters, 2010, 259(1): 89-96. |
| [26] | Alakomi H L. Lactic acid permeabilizes gram-negative bacteria by disrupting the outer membrane [J]. Applied and Environmental Microbiology, 2000, 66(5): 2001-2005. |
| [27] | Pan M, Qian C, Huo S, et al. Gut-derived lactic acid enhances tryptophan to 5-hydroxytryptamine in regulation of anxiety via Akkermansia muciniphila [J]. Gut Microbes, 2025, 17(1): 1-20. |
| [28] | Müller J, Radej J, Horak J, et al. Lactate: The Fallacy of Oversimplification [J]. Biomedicines, 2023, 11(12): 3192. |
| [29] | Zhang Y, Peng Q, Zheng J, et al. The function and mechanism of lactate and lactylation in tumor metabolism and microenvironment [J]. Genes & Diseases, 2023, 10(5): 2029-2037. |
| [30] | Yang X, Lu Y, Hang J, et al. Lactate-Modulated immunosuppression of Myeloid-Derived suppressor cells contributes to the radioresistance of pancreatic cancer [J]. Cancer Immunol Res. 2020, 8: 1440-51. |
| [31] | Peter K, Rehli M, Singer K, et al. Lactic acid delays the inflammatory response of human monocytes [J]. Biochemical & Biophysical Research Communications, 2015, 457(3): 412-418. |
| [32] | Huang X, Li Y, Fu M, et al. Polarizing macrophages in vitro [J]. Methods Mol Biol. 2018, 1784: 119-126. |
| [33] | Zhang J, Muri J, Fitzgerald G, et al. Endothelial lactate controls muscle regeneration from ischemia by inducing M2-like macrophage polarization [J]. Cell Metab. 2020, 31(6): 1136-153. |
| [34] | Selleri S, Bifsha P, Civini S, et al. Human mesenchymal stromal cell-secreted lactate induces M2-macrophage differentiation by metabolic reprogramming [J]. Oncotarget, 2016, 7(21): 30193-30210. |
| [35] | Colegio O, Chu NQ, Szabo A, et al. Functional polarization of tumour-associated macrophages by tumour-derived lactic acid [J]. Nature, 2014, 513, 559-563. |
| [36] | Lee Y S, Kim T Y, Kim Y, et al. Microbiota-derived lactate accelerates intestinal stem-cell-mediated epithelial development [J]. Cell Host & Microbe, 2018, 24(6): 833-846. |
| [37] | Allaire J M, Crowley S M, Law H T, et al. The intestinal epithelium: central coordinator of mucosal immunity [J]. Trends in Immunology, 2018, 39(9): 677-696. |
| [38] | Rodriguez-Colman M J, Schewe M, Meerlo M, et al. Interplay between metabolic identities in the intestinal crypt supports stem cell function [J]. Nature, 2017, 543(7645): 424-427. |
| [39] | Zhang D, Gao J, Zhu Z, et al. Lysinel-lactylation is the dominant lactylation isomer induced by glycolysis [J]. Nature Chemical Biology, 2025, 21(1): 91-99. |
| [40] | Li H, Liu C, Li R, et al. AARS1 and AARS2 sense l-lactate to regulate cGAS as global lysine lactyltransferases [J]. Nature, 2024, 634(8036): 1229-1237. |
| [41] | Moreno-Yruela C, Zhang D, Wei W, et al. Class I histone deacetylases (HDAC1-3) are histone lysine delactylases [J]. Science advances, 2022, 8(3): eabi6696. |
| [42] | Bao C, Ma Q, Ying X, et al. Histone lactylation in macrophage biology and disease: from plasticity regulation to therapeutic implications [J]. EBioMedicine, 2024, 111(000): 1-17. |
| [43] | Yue X, Zhou Q, Jia M, et al. Ulk1-mediated metabolic reprogramming regulates vps34 lipid kinase activity by its lactylation [J]. SSRN Electronic Journal, 2022, 9(22): 1-17. |
| [44] | Wei Y, Guo H, Chen S, et al. Regulation of macrophage activation by lactylation in lung disease [J]. Frontiers in Immunology, 2024, 15(15): 1427739. |
| [45] | 傅梦蝶, 廖伟. 组蛋白乳酸化修饰的研究进展[J]. 赣南医科大学学报, 2024, 44(11): 1181-1187. |
| [46] | Ju J, Zhang H, Lin M. The alanyl-tRNA synthetase AARS1 moonlights as a lactyltransferase to promote YAP signaling in gastric cancer [J]. J Clin Invest. 2024, 134(10): e174587. |
| [47] | Cai H, Chen X, Liu Y, et al. Lactate activates trained immunity by fueling the tricarboxylic acid cycle and regulating histone lactylation [J]. Nature Communications, 2025, 16(1): 3230. |
| [48] | Allis CD, Jenuwein T. The molecular hallmarks of epigenetic control [J]. Nat Rev Genet. 2016, 17(8): 487-500. |
| [49] | Chu X, Di C, Chang P, et al. Lactylated histone H3K18 as a potential biomarker for the diagnosis and predicting the severity of septic shock [J]. Front Immunol, 2021, 12: 786666. |
| [50] | Lu Z, Fang P, Li S, et al. Lactylation of Histone H3k18 and Egr1 Promotes Endothelial Glycocalyx Degradation in Sepsis‐Induced Acute Lung Injury [J]. Advanced Science, 2025, 12(7): 1-24. |
| [51] | Ziogas A, Novakovic B, Ventriglia L, et al. Long-term histone lactylation connects metabolic and epigenetic rewiring in innate immune memory [J]. Cell, 2025, 188(11): 2992-3012. |
| [52] | Nguyen N T B, Gevers S, Kok R N U, et al. Lactate controls cancer stemness and plasticity through epigenetic regulation [J]. Cell Metabolism, 2025, 37(4): 903-919. |
| [53] | Guan B, Zhou M, Dai W, et al. Peritumoral colonic epithelial cell-derived GDF15 sustains colorectal cancer via regulation of glycolysis and histone lactylation [J]. Nature Aging, 2025, 5(12): 2449-2465. |
| [54] | Dai J, Huang Y J, He X, et al. Acetylation Blocks cGAS Activity and Inhibits Self-DNA-Induced Autoimmunity [J]. Cell, 2019, 176(6): 1447-1460. |
| [55] | Yang K, Fan M, Wang X, et al. Lactate promotes macrophage HMGB1 lactylation, acetylation, and exosomal release in polymicrobial sepsis [J]. Cell Death Differ, 2022, 29(1): 133-146. |
APA Style
Hou, L., Sun, B., Sun, J., He, X., Deng, W., et al. (2026). Metabolic Function of Lactate and the Regulation and Biological Functions of Lactylation Modification. Science Research, 14(3), 62-70. https://doi.org/10.11648/j.sr.20261403.11
ACS Style
Hou, L.; Sun, B.; Sun, J.; He, X.; Deng, W., et al. Metabolic Function of Lactate and the Regulation and Biological Functions of Lactylation Modification. Sci. Res. 2026, 14(3), 62-70. doi: 10.11648/j.sr.20261403.11
@article{10.11648/j.sr.20261403.11,
author = {Linyue Hou and Baosheng Sun and Jinhui Sun and Xueqin He and Weixi Deng and Yuneng Yang},
title = {Metabolic Function of Lactate and the Regulation and Biological Functions of Lactylation Modification},
journal = {Science Research},
volume = {14},
number = {3},
pages = {62-70},
doi = {10.11648/j.sr.20261403.11},
url = {https://doi.org/10.11648/j.sr.20261403.11},
eprint = {https://article.sciencepublishinggroup.com/pdf/10.11648.j.sr.20261403.11},
abstract = {Lactate has long been regarded as a metabolic end product of glycolysis. However, recent studies have revealed its multiple functions in energy supply, signal transduction, and epigenetic regulation. This article systematically reviews the production, transport, and metabolic mechanisms of lactate, with an emphasis on its biological roles in immune regulation, inflammation control, intestinal homeostasis maintenance, and tumor microenvironment remodeling. In addition, it provides an in-depth discussion of the "writers-erasers-readers" regulatory network of lactylation. Lactylation is a lactate-driven post-translational modification that occurs on both histone and non-histone proteins. Histone lactylation primarily regulates gene transcription by altering chromatin conformation and plays key roles in macrophage polarization, inflammation resolution, and tumor progression. Non-histone lactylation extensively modifies various proteins, including metabolic enzymes and transcription factors, and participates in signal transduction, immune escape, and DNA repair. The two types of lactylation exhibit significant differences in modification sites, reaction mechanisms, and functional effects, together forming an important bridge between metabolism and epigenetic regulation. Current studies suggest that targeting lactate metabolism or lactylation-related regulatory pathways holds therapeutic potential in tumors, inflammatory diseases, and metabolic disorders. Future efforts are needed to further elucidate the dynamic regulatory networks of lactylation under pathological conditions and to facilitate its clinical translation.},
year = {2026}
}
TY - JOUR T1 - Metabolic Function of Lactate and the Regulation and Biological Functions of Lactylation Modification AU - Linyue Hou AU - Baosheng Sun AU - Jinhui Sun AU - Xueqin He AU - Weixi Deng AU - Yuneng Yang Y1 - 2026/05/29 PY - 2026 N1 - https://doi.org/10.11648/j.sr.20261403.11 DO - 10.11648/j.sr.20261403.11 T2 - Science Research JF - Science Research JO - Science Research SP - 62 EP - 70 PB - Science Publishing Group SN - 2329-0927 UR - https://doi.org/10.11648/j.sr.20261403.11 AB - Lactate has long been regarded as a metabolic end product of glycolysis. However, recent studies have revealed its multiple functions in energy supply, signal transduction, and epigenetic regulation. This article systematically reviews the production, transport, and metabolic mechanisms of lactate, with an emphasis on its biological roles in immune regulation, inflammation control, intestinal homeostasis maintenance, and tumor microenvironment remodeling. In addition, it provides an in-depth discussion of the "writers-erasers-readers" regulatory network of lactylation. Lactylation is a lactate-driven post-translational modification that occurs on both histone and non-histone proteins. Histone lactylation primarily regulates gene transcription by altering chromatin conformation and plays key roles in macrophage polarization, inflammation resolution, and tumor progression. Non-histone lactylation extensively modifies various proteins, including metabolic enzymes and transcription factors, and participates in signal transduction, immune escape, and DNA repair. The two types of lactylation exhibit significant differences in modification sites, reaction mechanisms, and functional effects, together forming an important bridge between metabolism and epigenetic regulation. Current studies suggest that targeting lactate metabolism or lactylation-related regulatory pathways holds therapeutic potential in tumors, inflammatory diseases, and metabolic disorders. Future efforts are needed to further elucidate the dynamic regulatory networks of lactylation under pathological conditions and to facilitate its clinical translation. VL - 14 IS - 3 ER -
Department of Modern Agriculture, Zunyi Vocational and Technical College, Zunyi, China
Department of Modern Agriculture, Zunyi Vocational and Technical College, Zunyi, China
Department of Modern Agriculture, Zunyi Vocational and Technical College, Zunyi, China
Department of Modern Agriculture, Zunyi Vocational and Technical College, Zunyi, China
Department of Modern Agriculture, Zunyi Vocational and Technical College, Zunyi, China
Department of Modern Agriculture, Zunyi Vocational and Technical College, Zunyi, China
Information