In order to identify flower-visiting insects on sesame plants and characterize the community structure, ecological survey was conducted in Bilone agroecological farm in 2022 and 2023, in 15 experimental plots (6x5.5 m each) each year, created in a 1,600 m² area. Insects were captured, stored in papillotes (Lepidoptera) or in vials containing 70° alcohol (other adults) and identified at the species level in laboratory. A total of 1,703 specimens were captured. They belonged to five orders, 12 families, 18 genera and 19 species. Hymenoptera was mostly collected order (91.5%) followed by Diptera (4.5%), Lepidoptera (1.8%), Neuroptera (0.9%) and Orthoptera (1.3%). Apidae was the most collected family (42.4%) followed by Formicidae (34.1%), Megachilidae (11.6%) while other families were rare: Acrididae (1.3%), Ascalapidae (0.9%), Calliphoridae (0.5%), Eumenidae (0.7%), Halictidae (2.2%), Muscidae (4.0%), Nymphalidae (1.3%), Pieridae (0.5%), and Vespidae (0.6%). Apis mellifera adansonii (Apidae: 30.6%) was the most recorded species, followed by Paratrechina longicornis (Formicidae: 12.3%), Pheidole megacephala (Formicidae: 9.4%), Myrmicanioa opaciventris (Formicidae: 8.9%), Megachile cincta (Megachilidae: 7.0%), Amegilla calens (Apidae: 6.2%), Xylocopa olivacea (Apidae: 5.6%), Megachile kamerunensis (Megachilidae: 4.6%), Musca domestica (Diptera: 4.0%), Camponotus maculatus (Formicidae: 3.65%), Lasioglossum hancocki (Halictidae: 2.2%), and Pteropera carnapi (Acrididae: 1.3%). Calliphora vicina (Calliphoridae) was recorded exclusively in 2022. Two exotic Diptera (Cl. vicina and Mu. domerstica) were myiasigenic species. The exotic Eumenidae Delta sp. and the afrotropical predator Ascalaphus africanus (Ascalapidae) were recorded as well as the phytophagous Acrididae Pe. carnapi. Potential pests (Nymphalidae, Pieridae and Acrididae) cumulatively represented 3.1% of the collection. The community was highly diversed and lowly dominated by a few species. Ca. maculatus was simply abundant in 2023. Amegilla calens, Ap. mellifera adansonii, Me. cincta, Me. kamerunensis, Mu. domestica, My. opaciventris, Pa. longicornis, Ph. megacephala and Xy. olivacea were simply abundant. Amegilla calens and Me. cincta, were co-dominants in 2022. Ca. maculatus and Me. kamerunensis were co-dominants in 2023. Apis mellifera adansonii, Pa. longicornis, Ph. megacephala, My. opaciventris and Xy. olivacea were co-dominants in each year. Ca. maculates and Cl. vicina were rare in 2022. Bicyclus dorothea (Nymphalidae), Delta sp. and La. hancocki were rare in 2023. Acraea acerata (Nymphalidae), Ascalaphus africanus (Ascalapidae), Catopsilia florella (Pieridae), Pteropera carnapi (Acrididae) and Synagris conuta (Vespidae) were rare. High value of Motomura constant (m=0.777 in 2022) and Preston constant (m=0.726 in 2023) suggested least evolved pioneer assemblages with species competition limited to the physical space. Overall, flower visiting insects exhibited a global positive net association.
| Published in | American Journal of Entomology (Volume 9, Issue 1) |
| DOI | 10.11648/j.aje.20250901.14 |
| Page(s) | 28-54 |
| Creative Commons |
This is an Open Access article, distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution and reproduction in any medium or format, provided the original work is properly cited. |
| Copyright |
Copyright © The Author(s), 2025. Published by Science Publishing Group |
Assemblage Composition, Co-Dominant Species, Rare Species, Theoretical Model, Assemblage Functioning, Sesame Plants
Orders / Families | Campaign | |||
|---|---|---|---|---|
2022 (%) | 2023 (%) | Total (%) | 2022 vs. 2023: Fisher’s exact test | |
Diptera Linnaeus, 1758 | ||||
Calliphoridae Hough (d), 1899 | 8 (0.5) | - | 8 (0.5) | χ²=8.607; df=1; p=0.008 * |
Muscidae Latreille, 1802 | 29 (1.7) | 39 (2.3) | 68 (4.0) | χ²=0.273; df=1; p=0.615 ns |
Total | 37 (2.2) | 39 (2.3) | 76 (4.5) | χ²=2.645; df=1; p=0.119 ns |
Hymenoptera Linnaeus, 1758 | ||||
Apidae Latreille, 1802 | 290 (17.0) | 431 (25.3) | 721 (42.3) | χ²=0.116; df=1; p=0.764 ns |
Eumenidae Leach, 1815 | - | 12 (0.7) | 12 (0.7) | χ²=9.326; df=1; p=0.003 * |
Formicidae Latreille, 1809 | 216 (12.7) | 364 (21.4) | 580 (34.1) | χ²=2.313; df=1; p=0.130 ns |
Halictidae Thomson, 1869 | - | 37 (2.23) | 37 (2.2) | χ²=34.026; df=1; p=9.6x10-9 * |
Megachilidae Latreille, 1802 | 97 (5.7) | 101 (5.9) | 198 (11.6) | χ²=7.878; df=1; p=0.005 * |
Vespidae Latreille, 1802 | 4 (0.2) | 7 (0.4) | 11 (0.6) | χ²=0.070; df=1; p=1.00 ns |
Total | 607 (35.6) | 952 (55.9) | 1,559 (91.5) | χ²=6.101; df=1; p=0.026 * |
Lepidoptera Linnaeus, 1758 | ||||
Nymphalidae Rafinesque, 1815 | 9 (0.5) | 13 (0.8) | 22 (1.3) | χ²=0.045; df=1; p=1.00 ns |
Pieridae Swainson, 1820 | 5 (0.3) | 3 (0.2) | 8 (0.5) | χ²=1.734; df=1; p=0.278 ns |
Total | 14 (0.8) | 16 (0.9) | 30 (1.8) | χ²=0.654; df=1; p=0.456 ns |
Neuroptera Linnaeus, 1758 | ||||
Ascalapidae Rambur, 1842 | 12 (0.7) | 4 (0.2) | 16 (0.9) | χ²=8.075; df=1; p=0.008 * |
Orthoptera Latreille, 1793 | ||||
Acrididae MacLeay, 1821 | 7 (0.4) | 15 (0.9) | 22 (1.3) | χ²=0.532; df=1; p=0.516 ns |
Global | 677 (39.8) | 1,026 (60.2) | 1,703 (100.0) | χ²=142.98; df=1; p=5.3x10-33 * |
Order/Family | Species name | Product | Origin, references | Campaign | ||
|---|---|---|---|---|---|---|
2022 (%) | 2023 (%) | Pooled (%) | ||||
Diptera | ||||||
Calliphoridae | Calliphora vicina Robineau-Desvoidy, 1830 | Nectar | NA, MS, a | 8 (0.5) | - | 8 (0.5) |
Muscidae | Musca domestica Linnaeus, 1758 | Nectar | ME, MS, b, c | 29 (1.7) | 39 (2.3) | 68 (4.0) |
Hymenoptera | ||||||
Apidae | Amegilla calens (Lepeletier De Saint-Fargeau, 1841) | Nectar, Pollen | AF, d, e | 59 (3.5) | 46 (2.7) | 105 (6.2) |
Apis mellifera adansonii Latreille, 1804 | Nectar | AF, e | 194 (11.4) | 327 (19.2) | 521 (30.6) | |
Xylocopa olivacea (Fabricius 1778) | Nectar, Pollen | AF, f | 37 (2.2) | 58 (3.4) | 95 (5.6) | |
Eumenidae | Delta sp. Saussure, 1855 | Predator | OW, Useful, g, k | - | 12 (0.7) | 12 (0.7) |
Formicidae | Camponotus maculatus (Fabricius, 1782) | Nectar | AF, h, i | 6 (0.4) | 53 (3.1) | 59 (3.5) |
Myrmicaria opaciventris Emery, 1893 | Nectar | AF, h, i | 80 (4.7) | 71 (4.2) | 151 (8.9) | |
Paratrechina longicornis (Latreille, 1802) | Nectar | AF, h, i | 90 (5.3) | 120 (7.0) | 210 (12.3) | |
Pheidole megacephala (Fabricius, 1793) | Nectar | AF, h, i, j | 40 (2.3) | 120 (7.0) | 160 (9.4) | |
Halictidae | Lasioglossum hancocki (Cockerell 1945) | Nectar, Pollen | AF, m | - | 37 (2.2) | 37 (2.2) |
Megachilidae | Megachile cincta (Fabricius, 1781) | Nectar | AF, l | 81 (4.8) | 38 (2.2) | 119 (7.0) |
Me. kamerunensis Friese, 1922 | Nectar, Pollen | AF, l | 16 (0.9) | 63 (3.7) | 79 (4.6) | |
Vespidae | Synagris conuta (Linnaeus, 1758) | Nectar, Pollen | AF, n, o | 4 (0.2) | 7 (0.04) | 11 (0.6) |
Lepidoptera | ||||||
Nymphalidae | Acraea acerata Hewitson, 1874 | Nectar | AF, Plant pest, p | 9 (0.5) | 6 (0.4) | 15 (0.9) |
Bicyclus dorothea (Cramer, 1779) | Nectar | AF, p, q, r | - | 7 (0.4) | 7 (0.4) | |
Pieridae | Catopsilia florella (Fabricius, 1775) | Nectar | AF, p, s | 5 (0.3) | 3 (0.2) | 8 (0.5) |
Neuroptera | ||||||
Ascalapidae | Ascalaphus africanus (McLachlan, 1871) | Predator | AF, Useful, t, u, v | 12 (0.7) | 4 (0.2) | 16 (0.9) |
Orthoptera | ||||||
Acrididae | Pteropera carnapi Ramme, 1929 | Phytophagous | AF, Plant pest, w | 7 (0.4) | 15 (0.9) | 22 (1.3) |
Total | 677 (39.8) | 1,026 (60.2) | 1,703(100.0) | |||
Statistical indices | Campaign | ||
|---|---|---|---|
I. 2022 | II. 2023 | III. Pooled years | |
A. Species richness indices | |||
Sample size n (%) | 677 (39.8) | 1,026 (60.2) | 1,703 (100.0) |
Observed species richness S | 16 | 18 | 19 |
Maximum abundance nmax | 194 | 327 | 521 |
Margalef’s index Mg | 2.301 | 2.452 | 2.419 |
Richness ratio d=S/n | 0.024 | 0.018 | 0.011 |
B. Non-parametric estimation of the "true" species richness | |||
ACE (SE=100*S/ACE) | 16 (100.0) | 18 (100.0) | 19 (100.0) |
ICE (SE=100*S/ICE) | 16 (100.0) | 18 (100.0) | 19 (100.0) |
Chao1 (SE=100*S/Chao1) | 16 (100.0) | 18 (100.0) | 19 (100.0) |
Chao2 (SE=(100*S/Chao2) | 16 (100.0) | 18 (100.0) | 19 (100.0) |
Jack.1 | 16 (100.0) | 18 (100.0) | 19 (100.0) |
Boostrap Mean | 16 (100.0) | 18 (100.0) | 19 (100.0) |
C. Species diversity indices | |||
Shannon-Weaver H’ | 2.232 | 2.294 | 2.329 |
Maximum Shannon-Weaver H’max=ln(S) | 2.773 | 2.890 | 2.944 |
Simpson index D | 0.145 | 0.150 | 0.143 |
D. Species dominance indices | |||
Berger-Parker dominance index IBP=nmax/n | 0.287 | 0.319 | 0.306 |
Hill’s first order diversity number N1=eH’ | 9.318 | 9.914 | 10.268 |
Hill’s second order diversity number N2=1/D | 6.878 | 6.667 | 6.983 |
Hill’s ratio: Hill=N2/N1 | 0.738 | 0.672 | 0.680 |
Estimated observed rare species: Chao1-N1 | 7 | 8 | 9 |
E. Evenness index | |||
Pielou’s index J=H’/H’max | 0.805 | 0.794 | 0.791 |
Comparison of the species diversity indices: I vs. II (Student t-test): | Shannon-Weaver index H’: t=-1.332; df=1,532.1; p=0.183 ns; Simpson index D: t=-0.433; df=1,635.1; p=0.665 ns; | ||
SAD theoretical model | Deviance; AIC (BIC) | ||
|---|---|---|---|
I. 2022 campaign 16 species; 677 specimens | II. 2023 campaign 18 species; 1,026 specimens | III. Pooled years 19 species; 1,703 specimens | |
McArthur’s Broken-Stick (BS) | 49.920; 129.097 (129.097) | 115.881; 209.095 (209.095) | 191.935; 298.518 (298.52) |
Motomura’s Log-linear (LL) | 23.191; 104.368 (105.140) * | 106.250; 201.464 (202.354) | 126.679; 235.261 (236.21) |
Preston’s Log-normal (LN) | 26.090; 109.267 (110.812) | 38.368; 135.582 (137.363) * | 53.367; 163.95 (165.84) * |
Zipf (Z) | 70.894; 154.071 (155.616) | 91.644; 188.857 (190.638) | 155.56; 266.142 (268.03) |
Zipf-Mandelbrot (ZM) | 21.740; 106.916 (109.234) | 79.141; 178.354 (181.025) | 103.945; 216.528 (219.36) |
Species 1/species 2 | tau τ | p-value | Species 1/species 2 | tau τ | p-value |
|---|---|---|---|---|---|
Acraea acerata | Apis mellifera adansonii | ||||
Ascalaphus africanus | 0.962 | 6x10-12 * | Me. cincta | -0.046 | 0.739 ns |
Bicyclus dorothea | 0.622 | 8x10-6 * | Me. kamerunensis | -0.070 | 0.617 ns |
Catopsilia florella | 0.593 | 2x10-5 * | My. opaciventris | -0.103 | 0.463 ns |
Pteropera carnapi | 0.843 | 2x10-9 * | Pa. longicornis | -0.056 | 0.689 ns |
Amegilla calens | Pe. carnapi | -0.082 | 0.559 ns | ||
Acraea acerata | 0.660 | 2x10-6 * | Ph. megacephala | -0.070 | 0.617 ns |
Apis mellifera adansonii | -0.135 | 0.335 ns | Sy. conuta | -0.103 | 0.462 ns |
As. africanus | 0.655 | 3x10-6 * | Xy. olivacea | 0.639 | 5x10-6 * |
Bi. dorothea | 0.426 | 0.002 * | As.africanus | ||
Camponotus maculatus | 0.648 | 3x10-6 * | Pe. carnapi | 0.786 | 2x10-8 * |
Ct. florella | 0.403 | 0.004 * | Bi. dorothea | ||
Delta sp. | 0.449 | 0.001 * | As. africanus | 0.596 | 2x10-5 * |
Lasioglossum (Ipomalictus) hancocki | 0.449 | 0.001 * | Ct. florella | 0.449 | 0.001 * |
Megachile (Chalicodoma) cincta | 0.565 | 5x10-5 * | Pe. carnapi | 0.758 | 6x10-8 * |
Me. (Chalicodoma) kamerunensis | 0.744 | 1x10-7 * | Camponotus maculatus | ||
Myrmicaria opaciventris | 0.958 | 7x10-12 * | Ac. acerata | 0.685 | 9x10-7 * |
Paratrechina longicornis | 0.775 | 3x10-8 * | As. africanus | 0.634 | 6x10-6 * |
Pe. carnapi | 0.570 | 5x10-5 * | Camponotus maculatus | ||
Pheidole megacephala | 0.759 | 5x10-8 * | Bi. dorothea | 0.634 | 6x10-6 * |
Synagris conuta | 0.545 | 9x10-5 * | Ct. florella | 0.206 | 0.139 ns |
Xy. olivacea | -0.015 | 0.915 ns | La. hancocki | 0.663 | 2x10-6 * |
Apis mellifera adansonii | Me. cincta | 0.399 | 0.004 * | ||
Ac. acerata | -0.197 | 0.158 ns | Me. kamerunensis | 0.885 | 2x10-10 * |
As. Africanus | -0.222 | 0.111 ns | My. opaciventris | 0.659 | 2x10-6 * |
Bi. dorothea | 0.049 | 0.723 ns | Pa. longicornis | 0.709 | 4x10-7 * |
Ca. maculatus | 0.036 | 0.798 ns | Pe. carnapi | 0.827 | 3x10-9 * |
Ct. florella | -0.263 | 0.059 ns | Ph. megacephala | 0.866 | 6x10-10 * |
Delta sp. | 0.066 | 0.637 ns | Sy. conuta | 0.465 | 0.001 * |
La. hancocki | 0.066 | 0.637 ns |
Species 1/species 2 | tau τ | p-value | Species 1/species 2 | tau τ | p-value |
|---|---|---|---|---|---|
Calliphora vicina | La. hancocki | ||||
Ac. acerata | 0.652 | 3x10- * | Ac. acerata | 0.652 | 3x10-6 * |
Am. calens | 0.472 | 0.001 * | As. africanus | 0.596 | 2x10-5 * |
Ap. mellifera adansonii | -0.329 | 0.018 * | Bi. dorothea | 0.959 | 6x10-12 * |
As. africanus | 0.715 | 3x10-7 * | Ct. florella | 0.408 | 0.003 * |
Bi. dorothea | -0.082 | 0.559 ns | Me. cincta | 0.449 | 0.001 * |
Ca. maculatus | 0.236 | 0.091 ns | La. hancocki | ||
Ct. florella | 0.490 | 5x10-4 * | Me. kamerunensis | 0.573 | 4x10-5 * |
Delta sp. | -0.082 | 0.559 ns | Pe. carnapi | 0.791 | 1x10-8 * |
La. hancocki | -0.082 | 0.559 ns | Sy. conuta | 0.746 | 9x10-8 * |
Me. cincta | 0.426 | 0.002 * | Me. cincta | ||
Me. kamerunensis | 0.520 | 2x10-4 * | Ac. acerata | 0.627 | 7x10-6 * |
Musca domestica | 0.359 | 0.010 * | As. africanus | 0.622 | 8x10-6 * |
My. opaciventris | 0.393 | 0.005 * | Bi. dorothea | 0.426 | 0.002 * |
Pa. longicornis | 0.474 | 0.001 * | Ct. florella | 0.403 | 0.004 * |
Ph. megacephala | 0.546 | 9x10-5 * | Me. kamerunensis | 0.518 | 2x10-4 * |
Pe. carnapi | 0.320 | 0.022 * | Pe. carnapi | 0.532 | 1x10-4 * |
Sy. comuta | 0.373 | 0.008 * | Sy. conuta | 0.545 | 9x10-5 * |
Xy. olivacea | -0.265 | 0.057 ns | Me. kamerunensis | ||
Ct. florella | Ac. acerata | 0.822 | 4x10-9 * | ||
As. africanus | 0.670 | 2x10-6 * | As. africanus | 0.797 | 1x10-8 * |
Pe. carnapi | 0.286 | 0.040 * | Bi. dorothea | 0.546 | 9x10-5 * |
Delta sp. | Ct. florella | 0.493 | 4x10-4 * | ||
Ac. acerata | 0.652 | 3x10-6 * | Pe. carnapi | 0.725 | 2x10-7 * |
As. africanus | 0.596 | 2x10-5 * | Sy. conuta | 0.653 | 3x10-6 * |
Bi. dorothea | 0.959 | 6x10-12 * | Mu. domestica | ||
Delta sp. | Ac. acerata | 0.648 | 4x10-6 * | ||
Ca. maculatus | 0.663 | 2x10-6 * | Am. calens | 0.758 | 6x10-8 * |
Ct. florella | 0.408 | 0.003 * | Ap. mellifera adansonii | -0.131 | 0.348 ns |
La. hancocki | 1.000 | 8x10-13 * | As. africanus | 0.609 | 1x10-5 * |
Me. cincta | 0.449 | 0.001 * | Bi. dorothea | 0.487 | 5x10-4 * |
Me. kamerunensis | 0.573 | 4x10-5 * | Ca. maculatus | 0.636 | 5x10-6 * |
My. opaciventris | 0.462 | 0.001 * | Ct. florella | 0.359 | 0.010 * |
Pa. longicornis | 0.439 | 0.002 * | Delta sp. | 0.510 | 3x10-4 * |
Pe. carnapi | 0.791 | 1x10-8 * | La. hancocki | 0.510 | 3x10-4 * |
Ph. megacephala | 0.573 | 4x10-5 * | Me. cincta | 0.346 | 0.013 * |
Sy. conuta | 0.746 | 9x10-8 * | Me. kamerunensis | 0.688 | 8x10-7 * |
Species 1/species 2 | tau τ | p-value | Species 1/species 2 | tau τ | p-value |
|---|---|---|---|---|---|
My. opaciventris | 0.734 | 1x10-7 * | Ph. megacephala | ||
Pa. longicornis | 0.702 | 5x10-7 * | Ac. acerata | 0.803 | 9x10-9 * |
Pe. carnapi | 0.602 | 2x10-5 * | As. africanus | 0.797 | 1x10-8 * |
Ph. megacephala | 0.673 | 1x10-6 * | Bi. dorothea | 0.546 | 9x10-5 * |
Sy. conuta | 0.529 | 2x10-4 * | Ct. florella | 0.520 | 2x10-4 * |
Xy. olivacea | 0.005 | 0.971 ns | La. hancocki | 0.573 | 4x10-5 * |
My. opaciventris | Me. cincta | 0.534 | 1x10-4 * | ||
Ac. acerata | 0.604 | 2x10-5 * | Me. kamerunensis | 0.983 | 2x10-12 * |
As. Africanus | 0.598 | 2x10-5 * | Pe. carnapi | 0.703 | 5x10-7 * |
Bi. dorothea | 0.439 | 0.002 * | Sy. conuta | 0.675 | 1x10-6 * |
Ct. florella | 0.393 | 0.005 * | Sy. conuta | ||
My. opaciventris | Ac. acerata | 0.751 | 8x10-8 * | ||
La. hancocki | 0.462 | 0.001 * | As. africanus | 0.804 | 8x10-9 * |
Me. cincta | 0.547 | 9x10-5 * | Bi. dorothea | 0.780 | 2x10-8 * |
Me. kamerunensis | 0.754 | 7x10-8 * | Ct. florella | 0.814 | 6x10-9 * |
Pa. longicornis | 0.784 | 2x10-8 * | Pe. carnapi | 0.573 | 4x10-5 * |
Pe. carnapi | 0.524 | 2x10-4 * | Xy. olivacea | ||
Ph. megacephala | 0.769 | 4x10-8 * | Ac. acerata | 0.039 | 0.783 ns |
Sy. conuta | 0.537 | 1x10-4 * | As. africanus | 0.013 | 0.926 ns |
Pa. longicornis | Bi. dorothea | 0.309 | 0.027 * | ||
Ac. acerata | 0.654 | 3x10-6 * | Ca. maculatus | 0.198 | 0.156 ns |
As. africanus | 0.649 | 3x10-6 * | Ct. florella | -0.009 | 0.949 ns |
Bi. dorothea | 0.416 | 0.003 * | Delta sp. | 0.327 | 0.019 * |
Ct. florella | 0.404 | 0.004 * | La. hancocki | 0.327 | 0.019 * |
La. hancocki | 0.439 | 0.002 * | Me. cincta | 0.205 | 0.143 ns |
Pa. longicornis | Me. kamerunensis | 0.098 | 0.482 ns | ||
Me. cincta | 0.534 | 1x10-4 * | My. opaciventris | 0.020 | 0.886 ns |
Me. kamerunensis | 0.814 | 6x10-9 * | Pa. longicornis | 0.115 | 0.410 ns |
Ph. megacephala | 0.829 | 3x10-9 * | Pe. carnapi | 0.160 | 0.250 ns |
Pe. carnapi | 0.562 | 6x10-5 * | Ph. megacephala | 0.098 | 0.482 ns |
Sy. conuta | 0.537 | 1x10-4 * | Xy. olivacea | ||
Sy. conuta | 0.140 | 0.317 ns |
Ac. acerata | Acraea acerata Hewitson. 1874 |
ACE | Abundance Coverage-based Estimator |
AIC | Akaike Information Criteria |
Am. calens | Amegilla calens (Lepeletier De Saint-Fargeau.1841) |
Ap. mellifera adansonii | Apis mellifera adansonii Latreille. 1804 |
As. africanus | Ascalaphus africanus (McLachlan. 1871) |
Bi. dorothea | Bicyclus dorothea (Cramer. 1779) |
BC | Bray-Curtis Index |
BIC | Bayesian Information Criteria |
BS | Broken-Stick Theoretical Model |
Ca. maculatus | Camponotus maculatus (Fabricius. 1782) |
Cl. vicina | Calliphora vicina Robineau-Desvoidy. 1830 |
Ct. florella | Catopsilia florella (Fabricius. 1775) |
CRC | Central Regional Council |
FAOSTAT | Food and Agricultural Organization Statistics |
GBIF | Global Biodiversity Information Facility |
GM | Geometric Theoretical Model |
IRAD/ARID | Institut de Recherche Agricole Pour le Développement/Agricultural Research Institute for Development |
ICE | Incidence Coverage-based Estimatorç |
ITIS | Integrated Taxonomic Information System |
OHIAM | Obala Higher Institute of Agriculture and Management |
La. hancocki | Lasioglossum (Ipomalictus) hancocki (Cockerell 1945) |
LL | Loglinear Theoretical Model |
LN | Lognormal Theoretical Model |
Me. kamerunensis | Megachile (Chalicodoma) kamerunensis Friese. 1922 |
Me. cincta | Megachile (Chalicodoma) cincta (Fabricius. 1781) |
Mu. domestica | Musca domestica Linnaeus. 1758 |
My. opacivenytrtis | Myrmicaria opaciventris Emery. 1893 |
Pa. longicornis | Paratrechina longicornis (Latreille. 1802) |
Pe. carnapi | Pteropera carnapi Ramme. 1929 |
Ph. megacephala | Pheidole megacephala (Fabricius. 1793) |
POWO | Plant of the World Online |
SAD | Species Abundance Distribution |
sp. | Undetermined Species |
Se. alatum | Sesamum alatum Thonn. |
Se. indicum | Sesamum indicum L. (1753) |
So. tuberosum | Solanum tuberosum L., 1753 |
SPSS | Statistical Package for the Social Sciences |
Sy. conuta | Synagris conuta (Linnaeus. 1758) |
VR | Variance Ratio |
Xy. olivacea | Xylocopa olivacea (Fabricius 1778) |
Z | Zipf Model |
ZM | Zipf-Mandelbrot Model |
| [1] | Hwang, L. S. Sesame oil. In Bailey’s industrial oil and fat products. 6th edition, Shahidu, F., Ed., New York: Wiley-Interscience; 2005, pp. 537-575. |
| [2] | Sene, B., Sarr, F., Diouf, D., Sow, M. S., Traore, Kane, A., Niang, M. Knowledge synthesis and acquired research of sesame (D. Sesamum indicum L.) in Senegal, International Journal of Biological and Chemical Sciences. 2018a, 12(3), 1469-1483. |
| [3] |
Integrated Taxonomic Information System. ITIS: Sesamum L. Taxonomic Serial No.: 34430. On-line database, www.itis.gov, CC0
https://doi.org/10.5066/F7KH0KBK (accessed 3 December 2024). |
| [4] |
Plant of the World Online. POWO: Sesamum alatum Thonn. Available from:
https://powo.science.kew.org/taxon/urn:lsid:ipni.org:names:675950-1 (accessed 3 December 2024). |
| [5] |
Bedigian, D. Sesamum radiatum Thonn. ex Hornem. Record from PROTA4U. Grubben, G. J. H. & Denton, O. A. (Eds.). PROTA (Plant Resources of Tropical Africa / Ressources végétales de l’Afrique tropicale), Wageningen, Netherlands.
http://www.prota4u.org/search.asp (accessed 3 December 2024). |
| [6] | Weiss, E. A. Oilseed Crops (second edition). United Kingdom: Blackwell Science LTD; 2000, pp. 1-355. |
| [7] | Söğüt, T. (2008). Effect of main and second cropping on seed yıeld, oil and protein content of Sesame (Sesamum indicum L.) Genotypes, Tukish Journal of Field Crops. 2008, 14(2), 64-71. |
| [8] |
Stevens, P. F. (2001 onwards). Angiosperm Phylogeny Website. Version 14, July 2017. Available from:
http://www.mobot.org/MOBOT/research/APweb/ (accessed 2 December 2024). |
| [9] | Kafiriti, E. M., Deckers, J. (2001). Sésame: Sesamum indicum L. Raemaekers, Agriculture en Afrique Tropicale. Bruxelles, Belgique: Direction Générale de la Coopération Internationale (DGCI); 2001, pp. 1-1634. |
| [10] | Andrade, P. B., Freitas, B. M., Rocha, E. E. M., Lima, J. A., Rufino, L. L. Floral Biology and pollination requirements of sesame (Sesamum indicum L.), Acta Scientiarum. Animal Sciences. Maringá. 2014, 36(1), 93-99, |
| [11] | Demol, J., Baudoin, J. P., Louant, B. P., Maréchal, R., Mergeai, G., Otoul, E. Amélioration des Plantes: Application aux Principales Espèces Cultivées en Régions. Tropicales. Gembloux: Les Presses Agronomiques de Gembloux; 2002, pp. 1-581. |
| [12] | Girmay, A. B. Sesame production, challenges and opportunities in Ethiopia, Agricultural Research & Technology. Open Access Journal. 2018, 15, 555972. |
| [13] | Saha, R. K., Dinar, M. A. M., Nabila, K. A., Roy, P. HPLC analysis and cell surface receptor binding activities of the crude aqueous and methanolic extract of Sesamum indicum, Asian Pacific Journal of Tropical Biomedicine. 2014, 4, S516-S520. |
| [14] | Bamigboye, A. Y., Okafor, A. C., Adepoju, O. T. Proximate and mineral composition of whole and dehulled Nigerian sesame seeds, African Journal of Food Science and Technology. 2010, 1(3), 71-75. |
| [15] |
Food and Agriculture Organization Corporate Statistical Database. FAOSTAT: Production: Crops and livestock products. Available from:
https://www.fao.org/faostat/fr/#data/QCL (accessed 5 December 2024). |
| [16] |
Sasu, D. D. Production quantity of sesame seeds in Africa 2021, by country. Available from;
https://www.statista.com/statistics/1298497/sesame-seeds-production-in-africa-by-country/ (accessed 4 December 2024). |
| [17] | Mohamed, E. A., Ahmed, H. I., Zaky, H. S., Badr, A. M. Sesame oil mitigates memory impairment, oxidative stress, and neurodegeneration in a rat model of Alzheimer's disease. A pivotal role of NF-κB/p38MAPK/BDNF/PPAR-γ pathways, Journal of Ethnopharmacology. 2021, 267, 113468, |
| [18] | Namiki, M. Nutraceutical functions of sesame: a review, Critical Reviews in Food Science and Nutrition. 2007, 7(47), 651-673. |
| [19] | Blal, A. E. H., Kamel, S. M., Mahfouz, H. M., Said, M. Impact of opened, non opened pollination and nitrogen fertilizer on sesame production in the reclaimed lands, Ismailia Governorate, Egypt, Cercetari Agronomice in Maldova. 2013, 46(3), 155. |
| [20] | Anilakumar, K. R., Pal, A., Khanum, F., Bawa, A. S. Nutritional, Medicinal and Industrial Uses of Sesame (Sesamum indicum L.) Seeds - An Overview, Agriculturae Conspectus Scientificus. 2010, 75(4), 159-168. |
| [21] | Amoo, S. O., Okorogbona, A. O. M., Du Plooy, C. P., Venter, S. L. Chapter 26 - Sesamum indicum. In Medicinal Spices and Vegetables from Africa, Kuete, V., Ed., Academic Press. pp. 549-579. |
| [22] | Mili, A., Das, S., Nandakumar, K., Lobo, R., (2021). A comprehensive review on Sesamum indicum L.: Botanical, ethnopharmacological, phytochemical, and pharmacological aspects, Journal of Ethnopharmacology. 2021, 281, 114503. |
| [23] | Ayouaz, S., Bensadia, D., Hamitri-Guerfi, F., Muhammad, D. R. A., Mouhoubi, K., Arab, R., Rahmani, Y., Guemouni, S., Hadjal, S., Madani, K. (2022). Impact of incorporating sesame oil (Sesamum indicum L.) in an Algerian frying oil and margarine formulation, North African Journal of Food and Nutrition Research. 2022, 6(14), 165-177. |
| [24] | Sene, B., Sarr, F., Diouf, D., Kane, A., Traore, D. Étude de la composition minérale et des teneurs en protéines et en matières grasses de huit variétés de sésame (Sesamum indicum L.) introduites au Sénégal pour un criblage variétal, Oilseeds & fats Crops and Lipids (OCL). 2018b, 2-7. |
| [25] | Kinman, M. L., Martin, J. A. Present status of sesame breeding in the United States, Agronomy Journal. 1954, 46, 24-27. |
| [26] | El Khier, M. K. S., Ishag, K. E. A., Yagoub, A. E. A. Chemical composition and oil characteristics of sesame seed cultivars grown in Sudan, Journal of Agricultural and Biological Science. 2008, 4(6), 761-766. |
| [27] | Alyemeni, M. N., Basahy, A. Y., Sher, H. (2011). Physico-chemical analysis and mineral composition of some sesame seeds (Sesamum indicum L.) grown in the Gizan area of Saudi Arabia, Journal of Medicinal Plants Research. 2011, 5(2), 270-274. |
| [28] | Borchani, C., Besbes, S., Blecker, C., Attia, H. (2010). Chemical characteristics and oxidative stability of sesame seed, sesame paste and olive oils, Journal of Agriculture, Science and Technology. 2010, 12, 585-596. |
| [29] | Jimoh, W. A., Fagbenro, O. A., Adeparusi, E. O. (2011). Effect of processing on some minerals, anti-nutrients and nutritional composition of sesame (Sesamum indicum) seed meals, Electronic Journal of Environmental, Agricultural and Food Chemistry. 2011, 10(1), 1864. |
| [30] | Hahm, T. S., Park, S. J., Martin Lo, Y. (2009). Effects of germination on chemical composition and functional properties of sesame (Sesamum indicum L.) seeds, Bioresource Technology. 2009, 100, 1643-1647. |
| [31] | Yogranjan, Satpute, G. K., Marabi, R. S., Mishra, M. K., Mishra, S. P. (2014). Global resurgence of sesame (Sesamum indicum L.) utilization: acurrent scenario, Indo-American Journal of Agricultural and Veterinary Sciences. 2014, 2(3), 2321-9602. |
| [32] | Kanu, P. J., Kerui, Z., Ming, Z. H., Haifeng, Q., Kanu, J. B., Kexue, Z. Sesame protein: Functional properties of sesame (Sesamum indicum L.) protein isolate as influenced by pH, temperature, time and ratio of flour to water during its production, Asian Journal of Biochemistry. 2007, 2, 289-301. |
| [33] | Nzikou, J. M., Mvoula-Tsiéri, M., Ndangui, C. B., Pambou-Tobi, N. P. G., Kimbonguila, A., Loumouamou, B., Silou, T., Desobry, S. Characterization of seeds and oil of sesame (Sesamum indicum L.) and the kinetics of degradation of the oil during heating, Research Journal of Applied Sciences, Engineering and Technology. 2010, 2(3), 227-232. |
| [34] | Dar, A. A., Arumugam, N. Lignans of sesame: purification methods, biological activities and biosynthesis - a review, Bioorganic Chemistry. 2013, 50, 1-10. |
| [35] | Prakash, K., Naik, S. N. Bioactive constituents as a potential agent in sesame for functional and nutritional application, Journal of Bioresource Engineering and Technology. 2014, 1, 48-66. |
| [36] | Prasad. Text book of field crop production. New Delhi: ICAR; 2002, pp. 1-821. |
| [37] | Gadade, B. V., Kachare, D. P., Satbhai, R. D., Naik, R. M. Nutritional composition and oil quality parameters of sesame (Sesamum indicum L.), Genotype. 2017, 3(7), 2454-8499. |
| [38] | Feng, B., Qian, K., Du, Y.-J. Floral Volatiles from Vigna un guiculata. Are Olfactory and Gustatory Stimulants for Ovipo sition by the Bean Pod Borer Moth Maruca vitrata. Insects. 2017, 8(2), 60. |
| [39] | Mohammadou, M., Fouelifack-Nintidem, B., Adamou, M., Taimanga, Kossini, D., Tsekane, S. J., Ngamaleu-Siewe, B., Kenne, E. L., Yomon, A. K., Kenne, M.. Diversity and Abun dance of Pest Insects Associated with Vigna unguiculata (L.) Walp, 1843 (Fabales: Fabaceae) in Bockle and Dang Localities (North-Cameroon). American Journal of Entomology. 2023, 7(2), 38-61. |
| [40] | Chhangani, G., Mahla, M. K., Swaminathan, R., Jain, H. K., Ahir, K. C. and Sharma, K. (2022). Diversity of Insect Fauna Associated with summer and Monsoon Cowpea [Vigna unguiculata (L.) Walp.]. Legume Research, 45 (6): 775-779. |
| [41] | Alam, M. Z., Crump, A. R., Haque, M. M., Islam, M. S., Hossain, E., Hasan, S. B., Hasan, S. B., & Hossain, M. S. (2016). Effects of Integrated Pest Management on Pest Damage and Yield Components in a Rice Agro-Ecosystem in the Barisal Region of Bangladesh. Frontiers in Environmental Science, 4: 22. |
| [42] | Tchuenguem Fohouo, F.-N. Activité de butinage et de pollinisation d’Apis mellifera adansonii Latreille (Hymenoptera: Apidae, Apinae) sur les fleurs de trois plantes à Ngaoundéré (Cameroun): Callistemon rigidus (Myrtaceae), Syzygium guineense var. macrocarpum (Myrtaceae) et Voacanga africana (Apocynaceae). Thèse de Doctorat d’Etat, Université de Yaoundé I, 2005. |
| [43] | Otiobo, A. E. N., Tchuenguem Fohouo, F.-N., Djieto-Lordon, C. Diversité de l’entomofaune floricole de Sesamum indicum (L.) 1753 (Pedaliaceae) et son impact sur les rendements fruitiers et grainiers à Bambui (Nord - Ouest, Cameroun), International Journal of Biological and Chemical Sciences. 2016, 10(1), 106-119. |
| [44] | Djonwangwé, D., Pando, J. B., Kameni, B. A. S., Bella, M. M. A., Tchuenguem, F. F.-N., Messi, J. (2017). Impact des activités de butinage de Xylocopa inconstans Smith F. 1874 (Hymenoptera: Apidae) et Megachile eurymera Smith 1864 (Hemenoptera: Megachilidae) sur la pollinisation et les rendements fruitier et grainier de Vigna unguiculata (L.) Walp. 1843 (Fabaceae) à Maroua, Extrême-Nord, Cameroun, Afrique Science. 2017, 13(5), 1-17. |
| [45] | Taïmanga, Mohammadou M., Mbianda P. A., Adamou M., Youssoufa O., Fouelifack-Nintidem B., Kenne Toukem A. S., Dabole O. M., Abdoul Aziz O., Tchoubou-Sale A., Yomon A. K., Tsekane S. J. & Kenne M. (2024). Impact of Plant Extracts on the Pollination Activity of Apis mellifera Linnaeus, 1758 (Hymenoptera: Apidae) on Flowers of Cowpea Variety Feekem, in Dang (Adamaoua, Cameroon). American Journal of Entomology, 8(2): 31-59. |
| [46] | Kengni, B. S., Ngakou, A., Tchuenguem Fohouo, F.-N.. Pollina tion and yield attributes of (cowpea) Vigna unguiculata L. Walp. (Fabaceae) as influenced by the foraging activity of Xylocopa ol ivacea Fabricius (Hymenoptera: Apidae) and inoculation with Rhizobium in Ngaoundere, Cameroon. International Journal of Agronomy and Agricultural Research. 2015, 6(2), 62-76. |
| [47] | Djidjonri, F. P., Nukenine, E. N., Koehler, H. H. Abundance and Diversity of Insect Pests on Maize, Cowpea and Okra in a Comparative Experiment Testing Effects of Intercropping and Insecticide in the Cameroonian Guinean Savannah and Sudano Sahelian Agro-ecological Zones, Journal of Experimental Agriculture International. 2019, 29(6), 1-20. |
| [48] | Mazi, S., Adamou, M. , Issaya Issaya, K., Mamoudou, J., Faïbawa E. (2020) Impact of Amegilla calens and Apis mellifera Pollination on Gossypium hirsutum var. QR1302 Flowers at Tchabbal-Mounguel (Ngaoundéré, Cameroon). Open Journal of Ecology, 2020, 10, 445-459. |
| [49] | Manfo, D. A. Pratiques et enjeux de l’agroforesterie dans la zone de contact forêt-savane: le cas d’Obala dans la Région du Centre au Cameroun, Revue Scientifique et Technique Forêt et Environnement du Bassin du Congo. 2018, 11, 66-78. |
| [50] | Manfo, D. A., Tchindjang, M., Youta, H. J. Systèmes agroforestiers et conservation de la biodiversité dans un milieu fortement anthropisé: le cas d’Obala, Revue Scientifique et Technique Forêt et Environnement du Bassin du Congo. 2015, 5, 22-34. |
| [51] | Moudelsia, O. B., Otiobo Atibita, N. E., Heumou, C. R., Pando, J. B., Djonwangwe, D. Foraging behavior of three wild bees (Amegilla sp., Sphecodes albilabris and Lasioglossum albipes) on male flowers of Zea mays (Poaceae) at Maroua, Cameroon. European Scientific Journal, ESJ. 2022, 18(27), 12. |
| [52] | Pharaon Mbianda, A., Douka, C., Dounia, Eloundou, C. E., Tchuen guem Fohouo. F.-N. Pollination efficiency of Apis mellifera L. (Hymenoptera: Apidae) on flowers of Vigna unguiculata (L.) Walp. (Fabaceae) at Bilone (Obala, Cameroon). International Jopurnal of Bioscience. 2019, 14(1): 1-11. |
| [53] | Adamou, M., Nepide, N. C., Mazi, S., Yatahaï, C. M. Impact of the pollinating activity of Apis mellifera (Hymenoptera: Apidae) on pod and seed yields of Vigna unguiculata (Faba ceae) variety BR1 in Djoumassi (North Cameroon). Cameroon Journal of Biological and Biochemical Sciences. 2020, 28(2), 146-159. |
| [54] | Pauly A., Breat, Y., Tchibozo, S., Aikpe, C. & Boeve, J. L. Hymenoptera Apoidea et Braconidae de quelques forêts sacrées du Sud-Bénin. Bulletin de la Société Royale Belge d'Entomologie/Bulletin van de Koninklijke Belgische Vereniging voor Entomologie. 2009, 45, 121-129. |
| [55] | Ige, O. E., Olotuah, O. F., Akerele, V. Floral biology and pol lination ecology of cowpea (Vigna unguiculata L. Walp). Modern Applied Science. 2011, 5(4): 74-82. |
| [56] | Kwapong, P. K., Danquah, P. O. A., Asare, A. T. Insect floral visitors of cowpea (Vigna unguiculata L.). Annals of Biologi cal Research. 2013, 4, 12-18. |
| [57] | Manap, T., Fajri, M. Diversity of Bees and Wasp (Hymenoptera) in Cowpea (Vigna sinensis L.) in Agricultural Area at Martapura District, Banjar Regency, South Kalimantan, Journal of Science and Technology. 2020, 9, 29-33. |
| [58] | Sandeep Kumar J., Rex B,, Irulandi S., Prabhu S. A Review on Diversity, Bio-Ecology, Floral Resources and Behavior of Blue Banded Bees. International Journal of Current Microbiology and Applied Sciences. 2019, 8(07), 580-587. |
| [59] | Atmowidi T., Nur Cholis M., Maulana A., Priawandiputra W., Kahono S. Effectiveness of Pollinator Insects in Increasing Fruit Formation of Pummelo (Citrus maxima (Burm.) Merr.). Australian Journal of Crop Science. 2022, 16(09), 1078-1083 |
| [60] | Hussain K., Attaullah M., Rafi M. A., Khan H., Waris A., Zeb A., Baset A. Diversity of Carpenter Bee Fauna (Xylocopa spp.) in Dir Lower, Khybfr Pakhtunkhwa, Pakistan. Indo American Journal of Pharmaceutical Sciences. 019, 06(10), 13512-13520. |
| [61] | Kameli S. M., Blal A. E. H., Mahfouz H. M., Said M. Pollinator Fauna of Sesame Crop (Ssesamum indicum L) in Ismaila Governorate, Egypt. Cercetări Agronomice în Moldova. 2013, XLVI, 2(154), 53-64. |
| [62] | Ajao A. M., Oladimeji Y. U., Aderolu I. Survey of Crop-Plants and Honey Bee Pollinazation: A Stimulus to Food Security in Kwara State Nigeria. Journal of Agriculture and Environment. 2018, 14(1), 123-134. |
| [63] | Taimanga, Adamou M., Tchindebe G., Mohammadou M., Youssoufa O., Fouelifack-Nintidem B., Tchiaze Ifoue A. V., Kenne Toukem A. S., Massah Dabole O., Abdoul Aziz O., Tchoubou-Sale A., Tsekane S. J., Kosini D., Pharaon Mbianda A., Kenne M. Effect of the Botanical Insecticides on Amegilla Friese, 1897 (Hymenoptera: Apidae) Foraging on the Cowpea Flowers in Dang (Adamaoua, North-Cameroon). American Journal of Entomology. 2024, 8(3), 76-101. |
| [64] | Tchuenguem Fohouo, F.-N., Ngakou, A., Kengni, B. S. Polli nation and yield responses of cowpea (Vigna unguiculata L. Walp.) to the foraging activity of Apis mellifera adansonii (Hymenoptera: Apidae) at Ngaoundéré (Cameroon). African Journal of Biotechnology. 2009, 8(9), 1988-1996. |
| [65] | Zahra, T., Mehran, R., Asghar, T. A. Cowpea: Insect Pest Management. In Agricultural Research Updates. Volume 26, Prathamesh, G., Srushti, M., Eds., Nova Science Pulishers, Inc.; 2019, pp. 1-48. |
| [66] | Fouelifack-Nintidem, B., Yetchom-Fondjo, J. A., Tsekane, S. J., Ngamaleu-Siewe, B., Kenne, E. L., Biawa Kagmegni, M., Tuekam-Kowa, P. S., Yomon, A. K., Kentsop-Tsafong, R. M., Dim-Mbianda, A. M., Kenne, M. Diversity and abundance of pest insects associated with the Ethiopian eggplant plants Solanum aethiopicum Linnaeus, 1756 (Solanaceae) in Balessing (West-Cameroon), American Journal of Entomology. 2021, 5(3), 70-91. |
| [67] | Ngamaleu-Sewe, B., Fouelifack-Nintidem, B., Yetchom-Fondjo, J. A., Moumite Mohamed, B., Tsekane, S. J., Kenne, E. L., Biawa-Kagmegni, M., Tuekam Kowa, P. S., Fantio, R. M., Yomon, A. K., Kenne, M., 2021. Abundance and diversity of insects associated with Solanum tuberosum L. 1753 (Solanaceae) after insecticide treatments in Balessing (West-Cameroon), American Journal of Entomology. 2021, 5(3), 51-69. |
| [68] | Klin-Khamer, P. G. L., De Jong, T. J. (1993). Attractiveness to pollinators: a plant’s dilemma, Oikos. 1993, 66, 180-184. |
| [69] |
Central Regional Council. CRC: Presentation of the Central Region. Available from:
https://region-centre.cm/?p=483 (accessed 11 December 2024). |
| [70] | Djimarbeye, C., Messi, P., Biye, E.-H. Household Waste Management and Environmental Risks in Obala, Central Region Cameroon, European Scientific Journal. 2024, 28, 516. |
| [71] | Kottek, M., Grieser, J., Beck, C., Rudolf, B., Rubel F. World Map of the Köppen-Geiger Climate Classification Updated, Meteorologische Zeitschrift. 2006, 15(3), 259-263. |
| [72] | Delvare, G., Arbelenc, H. P. 1989. Les insectes d’Afrique et d’Amérique tropicale: clés pour la reconnaissance des familles. Montpellier, France: CIRAD; 1989, pp 1-297. |
| [73] | Borror, D. J., White, R. E. Les insectes de l’Amérique du Nord (au nord du Mexique). Laprairie: Broquet, pp. 1-408. |
| [74] |
Eardley, C. D., Kuhlmann, M., Pauly A. Les genres et sous-genres d’abeilles de l’Afrique subsaharienne. Coopération Belge au Développement, ABC Taxa 9; 2010, pp. 1-152.
https://biblio.naturalsciences.be/rbins-publications/abc-txa/abc-taxa-09/abctaxa_vol9lr.pdf |
| [75] | Lecoq, M. Taxonomie et systématique des acridiens et principales espèces d’Afrique de l’Ouest. Montpellier, France: CIRAD. UPR Acridologie; 2010, pp. 1-106. |
| [76] | Brailovsky, H. Illustrated key for identification of the species included in the genus Leptoglossus (Hemiptera: Heteroptera: Coreidae: Coreini: Anisoscelini) and descriptions of five new species and new synonyms, Zootaxa. 2014, 3794, 143-178. |
| [77] | Tronquet, M. Catalogue des Coléoptères de France. Perpignan, France: Association Roussillonnaise d’Entomologie (ARE), Supplément au Tome XXIII-R; 2014, pp. 1-58. ISBN: 1288-5509. |
| [78] |
Taylor, B. The Ants of (sub-Saharan) Africa (Hymenoptera: Formicidae). Profusely illustrated with original drawings and photographs and Catalogue notes on all ant species described from sub-Saharan Africa. 2015. Available from:
https://antsofafrica.org/ (accessed 6 December 2024). |
| [79] | Zettler, J. A., Mateer, S. C., Link-Pérez, M. A., Bailey, J., Demars, G., Ness, T. To Key or Not to Key: A New Key to Simplify and Improve the Accuracy of Insect Identification, The American Biology Teacher. 2016, 78(8), 626-633. |
| [80] | Nihei, S. S., De Carvalho, C. J. B. The Muscini flies of the world (Diptera. Muscidae): identification key and generic diagnoses, Zootaxa. 2009, 1976, 1-24. |
| [81] | Whitworth, T. Keys to the genera and species of blow flies (Diptera: Calliphoridae) of the West Indies and description of a new species of Lucilia Robineau-Desvoidy, Zootaxa. 2010, 2663, 1-35. |
| [82] | Hewitt, C. G. The House-Fly: Musca domestica Linn: Its Structure. Habits. Development. Relation to Disease and Control. Cambridge University Press; 2011, pp. 5-6. ISBN: 978-0-521-23299-9. |
| [83] | Hölldobler, B., Wilson, E. O. The ants. Cambridge: Harvard University Press, Springer; 1990, pp. 1-732. |
| [84] | Carpenter, J. M. Review of Hawaiian Vespidae (Hymenoptera), Bishop Museum Occasional Papers. 2008, 99, 1-18. |
| [85] | Gusenleitner, J., Madl, M. Notes on Eumeninae (Insecta: Hymenoptera: Vespidae) of the Ethiopian Region based on the material of the Naturhistorisches Museum Wien (Austria), Annalen des Naturhistorischen Museums in Wien. B. 2012, 114, 9-25. |
| [86] | Wetterer, J. D. Worldwide spread of the African big-headed ant. Pheidole megacephala (Hymenoptera: Formicidae), Myrmecological News. 2012, 17, 51-62. |
| [87] | Pauly, A. The genus Chalicodoma in West and Central Africa. 2015. Available from: |
| [88] | Pauly, A. Ipomalictus Pauly. 1999. sous-genre de Lasioglossum nouveau pour la Région Orientale. avec des notes sur quelques espèces afrotropicales (Hymenoptera: Halictidae), Bulletin de l’Institut royal des Sciences naturelles de Belgique. Entomologie. 2001, 71, 145-154. |
| [89] | Oyerinde, A. A. Morphometric and wing landmarks analysis of races of Apis mellifera adansonii L. in Nigeria, Journal of Entomology and Zoology Studies. 2017, 5(5), 1374-1380. |
| [90] | Madl, M. A catalogue of the Apidae (Hymenoptera. Apoidea) of Eritrea, Linzer Biologische Beiträge. 2020, 52(1), 337-363. |
| [91] |
Global Biodiversity Information Facility. GBIF Secretariat: Eumenidae. GBIF Backbone Taxonomy. Checklist dataset. 2023a. Available from:
https://doi.org/10.15468/39omei (accessed via GBIF.org 6 November 2024). |
| [92] | Bivar-de-Sousa, A., Vasconcelos, S., Mendes, L. F., Larsen, T. B., Baker, J., Guilherme, J. L. Butterflies of Guinea-Bissau: VIII. New data, new reports, corrections and biodiversity (Lepidoptera: Papilionoidea). Zootaxa. 2016, 4201(1), 001-077. |
| [93] | Selis, M. Taxonomic revision of the subgenus Synagris (Paragris) de Saussure (Hymenoptera: Vespidae: Eumeninae), Journal of Natural History. 2023, 57(41-44), 1949-1962. |
| [94] | Aduse-Poku, K., Brattström, O., Kodandaramaiah, U., Lees, D. C., Brakefield, P. M., Wahlberg, N. (2015). Systematics and historical biogeography of the old world butterfly subtribe Mycalesina (Lepidoptera: Nymphalidae: Satyrinae), BMC Evolutionary Biology. 2015, 15, 167. |
| [95] |
Pauly, A. Les Xylocopa d'Afrique. Atlas Hymenoptera. 2016. Available from:
http://www.atlashymenoptera.net/page.aspx??id=84 (accessed 14 Bovember 2024). |
| [96] | Dongmo, M. A. K., Bonebrake, T. C., Fomena, A., Hanna, R. Life history notes on Bicyclus dorothea Cramer (Nymphalidea: Satyrinae) in Cameroon, Tropical Lepidoptera Research. 2017, 27(1), 28-32. |
| [97] | Kioko, E. N., Kochey, J. K., Mutua, M. N., Mwinzi, D. K. Field guide to the butterflies of the Sacred Mijikenda Kaya Forests World Herityage Site: Kaya Kauma Forest. Kenya: National Museum of Kenya; 2021, pp. 1-84. |
| [98] | Tjeder, B., Hansson, C. The Ascalaphidae of the Afrotropical Region (Neuroptera). 2. Revision of the tribe Ascalaphini (subfam. Ascalaphinae) excluding the genus Ascalaphus Fabricius, Entomologica Scandinavica Supplement. 1992, 41, 173-237. |
| [99] | Prost, A. The genus Ascalaphus (Fabricius. 1775) (Neuroptera. Ascalaphidae) in Africa. In Proceedings of the XIth International Symposium of Neuropterology, de Freiras Martins, A. M., Ventura, M., Eds., Açoreana Supplement 9. Sociedade Afonso Chaves: Ponta Delgada, Portugal; 2013, pp. 57-72. |
| [100] |
Oswald, J. D. Neuropterida Species of the World. A Catalogue and Monograph of the Species and Subspecies of the Extant and Extinct Neuroptera, Megaloptera, Raphidioptera and Glosselytrodea (Insecta: Neuropterida) of the World. Lacewing Digital Library. Research Publication No. 1. 2024. Available from:
http://lacewing.tamu.edu/SpeciesCatalog/Main (accessed 21 November 2024). |
| [101] | Yetchom Fondjo, J. A., Nzoko Fiemapong, A. R., Tindo, M., Duressa, T. F., Ivković, S., Husemann, M. (2024). Taxonomic review of the grasshopper genus Pteropera Karsch. 1891 (Orthoptera. Acrididea. Catantopinae) with description of three new species and a preliminary phylogeny of the Cameroonian species, Zookeys. 2024, 1216, 219-264. |
| [102] | Hammer, Ø., Harper, D. A. T., Ryan, P. D. (2001). PAST: Paleontological Statistics Software Package for Education and Data Analysis, Palaeontologia Electronica. 2001, 4(1), 1-9. |
| [103] |
Colwell, R. K. EstimateS: Statistical estimation of species richness and shared species from samples. Version 9. User's Guide and application. 2013. Available from:
http://purl.oclc.org/estimates (accessed 6 December 2024). |
| [104] | Schluter, D. A. (1984). A variance test for detecting species associations. with some example applications, Ecology. 1984, 65(3), 998-1005. |
| [105] | Chao, A., Chadzon, R. L., Colwell, R. K., Shen, T.-J. A new statistical approach for assessing similarity of species composition with incidence and abundance data, Ecology Letters. 2005, 8, 148-159. |
| [106] | McGill, B. J., Etienne, R. S., Gray, J. S., Alonso, D., Anderson, M. J., Benecha, H. K., Dornelas, M., Enquist, B. J., Green, J. L., He, F., Hurlbet, A. H., Magurran, A. E., Marquet, P. A., Maurer, B. A., Ostling, A., Soykan, C. U., Ugland, K. I., White, E. P. Species abundance distributions: moving beyond single prediction theories to integration within an ecological framework, Ecology Letters. 2007, 10(10), 995-1015. |
| [107] | Johnson, J. B., Omland, K. S. Model selection in ecology and evolution, Trends in Ecology & Evolution. 2004, 19(2), 101-108. |
| [108] | Li, W. Zipf's Law Everywhere, Glottometrics. 2002, 5, 14-21. |
| [109] | Koplenig, A. Using the parameters of the Zipf-Mandelbrot law to measure diachronic lexical. Syntactical and stylistic changes - a large-scale corpus analysis. Degruyter Mouton, Corpus Linguistics and Lingustic Theory. 2015, 1-34. |
| [110] | Le, D.-H., Pham, C.-K., Nguyen, T. T. T., and Bui, T. T. Parameter extraction and optimization using Levenberg-Marquardt algorithm. In Proceedings of 2012 IEEE conference. Fourth International Conference on Communications and Electronics (ICCE), Hanoi University of Science and Technology: Hanoi, Vietnam; 2012, pp. 434-437. |
| [111] | Murthy, Z. V. P. Nonlinear Regression: Levenberg-Marquardt Method. In Encyclopedia of Membranes, Drioli, E., Giorno, L., Eds., Berlin. Heidelberg Springer-Verlag.; 2014, pp. 1-3. |
| [112] | Latif, M. A., Rahman, M. M., Islam, M. R., Nuruddin, M. M. Survey of Arthropod Biodiversity in the Brinjal Field, Journal of Entomology. 2009, 6(1), 28-34. |
| [113] | Uno, S., Cotton, J., Philpott, S. M. Diversity, abundance, and species composition of ants in urban green spaces, Urban Ecosystems. 2010, 13, 425-441. |
| [114] | Akunne, C. E., Ononye, B. U., Mogbo, T. C. Insects: Friends or Enemies?, Global Journal of Biology, Agriculture & Health Sciences. 2013, 2(3), 134-140. |
| [115] | Adja, N. A., Danho, M., Alabi, T. A. F., Gnago, A. J., Zimmer, J. Y., Francis, F., Kouassi, P., Baudoin, J. P., Zoro Bi, I. A. Entomofauna associated with African oleaginous cucurbits (Lagenaria siceraria Molina (Standl. 1930) and Citrullus lanatus Thumb (Matsum & Nakai 1916)) and impact of pests on production, International Journal of Entomology. 2014, 50(3-4), 301-310. |
| [116] | Nghia, N. T., Srivastava, P. (2015). Biodiversity of beneficial insect associated with cowpea at Pantnagar - Uttarakhand - India, OMONRICE. 2015, 20, 73-79. |
| [117] | Obodji, A., Aboua, L. R. N., Tano, D. K. C., Seri-Kouassi, B. P. (2016). Inventory of entomofaune associated with African eggplant (Solanum aethiopicum L.) according to the phonological stages assessment of damages caused by insect pests, Journal of Advanced Studies in Agricultural, Biological and Environmental Sciences. 2016, 3(2), 2455-0221. |
| [118] | Babar, H. C., Asif, H. C., Abdul, G. L., Aslam, B., Imtaiz, A. N., Ammara, R., Fida, H. M., Mehroz, K., Farukh, A., Zehua Z. (2019). Insect Biodiversity in Brinjal Agro-Ecosystem, Pakistan Journal of Scientific and Industrial Research Series A: Physical Sciences. 2019, 62B (3), 199-205. |
| [119] | Chhangani, G., Mahla, M. K., Swaminathan, R., Jain, H. K., Ahir, K. C., Sharma, K. Diversity of Insect Fauna Associated with Summer and Monsoon Cowpea [Vigna unguiculata (L.) Walp.], Legume Research. 2022, 45(6), 775-779. |
| [120] | Mahmoud, M. F. Insects Associated with Sesame (Sesamun indicum L.) and the Impact of Insect Pollinators on Crop Production, Pesticides and Phytomedicine (Belgrade). 2012, 27(2), 117-129. |
| [121] | Padhy, D., Dash, L., Ramalxmi, V., Kumar Sahu, A. Insects Associated with Sesamum (Sesamum indicum L.) near Paralakhemundi region, International Journal of Bioinformatics and Biological Sciences. 2020, 8(Special Issue), 69-72. |
| [122] | Manawadu, D., Sharah, H. Review of Insects associated with Cowpea: the case of Maiduguri in the Semi-arid zone of Nigeria, Nigerian Journal of Entomology. 1990, 11, 100-111. |
| [123] | Zaki, A. Y., Aly, A. I. Biodiversity of Spider and Other Arthropods Inhabiting Cowpea under Effect of Fish Culture Water and Nitrogen Fertilization and its effect on Yield and Protein at Fayoum Governorate, Egypt, Acarines: Journal of the Egyptian Society of Acarology. 2018, 12, 87-98. |
| [124] | Rao, K. S., Kumar, Y., Yadav, S., Rawal, R. Diversity and Abundance of flower visiting insects associated with Sesame, Indian Journal of Entomology. 2022, 84(1), 77-81. |
| [125] | Morris, J. B. (). Food, industrial, intracentrial, and pharmaceutical uses of sesame genetic resources. In Trends in new crops and new uses, Janick, J. Whipkey, A, Eds., Arlington, VA, ASHS Press; 2002, pp. 153-156. |
| [126] |
Sheahan, C. M. Plant guides for sesame (Sesamum orientale) USDA-National Resources Conservation Service, Cape May Plant Materials Center, Cape May, NJ. 2014. Available from:
https://www.feedipedia.org/node/21661 (accessed 6 December 2024). |
| [127] | Langham, D. R. Growth and development of Sesame. USA: American Sesame Growers Association (ASGA), US Grown Sesame; 2008, pp. 1-45. |
| [128] | Ahohuendo, B. C., Sinebou, V. C. O., Ahoton, L. E., Etèka, A. C., Dansi, A., Ahanchédé, A., Hounhouigan, J. D., Sanni, A., de Foucault, B. Study of the phenology and floral biology of Sesamum radiatum Schumach. & Thon., a traditional leafy vegetable in domestication in Benin, Acta Botanica Gallica: Botany Letters. 2012, 159(3), 335-344. |
| [129] | Mahfouz, H. M., Kamel, S. M., Belal, A. H., Said, M. Pollinators visiting sesame (Sesamum indicum L.) seed crop with reference to foraging activity of some bee species. Cercetări Agronomice în Moldova, XLV; 2012, pp. 1-150. |
| [130] | Webster, A. D. Production systems and agronomy: Orchard Crops. In Encyclopedia of Applied Plant Sciences, Brian T. B., Ed. Elsevier; 2003, pp. 924-934 |
| [131] | Solar, R. R. C., Barlow, J., Andersen, A. N., Schoereder, J. H., Berenguer, E., Ferreira, J. N., Gardner, T. A. Biodiversity consequences of land-use change and forest disturbance in the Amazon: A multi-scale assessment using ant communities, Biological Conservation. 2016, 197, 98-107. |
| [132] | Biawa-Kagmegni, M., Foguieng-Saha, A. D., Guetsop-Ngouadjie, R. P., Tsekane, S. J., Fouelifack-Nintidem, B. Moumite Mohamed, B., Yetchom-Fondjo, J. A., Ngamaleu-Siewe, B., Kenne, E. L., Tuekam Kowa, P. S., Fantio, R. M., Yomon, A. K., Mbenoun Masse, P. S., Kenne, M., Fomena, A. Ants community structure in the urban and the city suburbs areas of Douala (Littoral-Cameroon), Journal of Insect Biodiversity. 2021, 025(2), 033-059. |
| [133] | Dzokou, V. J., Lontchi Fofe, N., Kamgaing Kouam, B. H., Yaouba, A., Tamesse, J. L. Fauna Pests Infesting Pepper (Piper nigrum L.) in Penja-Cameroon, American Journal of Entomology. 2021, 5(2), 32-38. |
| [134] | Carpentier, P., Leprêtre, A. Robustesse de quelques indices de diversité à l’échantillon, Océanis. 1999, 25(3), 435-455. |
| [135] | Iganaki, H. Mise au point de la loi de Motomura et essai d’une écologie évolutive, Vie Milieu. 1967, 18, 153-166. |
| [136] | Cagniant, H. Essai d'application de quelques indices et modèles de distributions d'abondances a trois peuplements de fourmis terricoles, Orsis. 1989, 4, 113-124. |
| [137] |
Errouissi, F., Jay-Robert, P., Lumaret, J.-P., Piau, A. Composition and Structure of Dung Beetle (Coleoptera: Aphodiidae, Geotrupidae, Scarabaeidae) Assemblages in Mountain Grasslands of the Southern Alps, Annals of the Entomological Society of America. 2004, 97(4), 701-709.
https://doi.org/10.1603/0013-8746(2004)097 [0701:CASODB] 2.0.CO; 2à. |
| [138] | Trouillet, J., Vattier-Bernard, G.. Phlébotomes du Mayombe Congolais (Diptera, Psychodidae). Analyse structurale de trois peuplements, Annales de Parasitologie Humaine et Comparée (Paris). 1983, 58(3), 291-300. |
| [139] | Komonen, A. & Elo, M. Ecological response hides behind the species abundance distribution: Community response to low-intensity disturbance in managed grasslands, Ecology and Evolution. 2017, 7, 8558-8566. |
| [140] | Yetchom-Fondjo, J. A., Kekeunou, S., Kenne, M., Missoup, A. D., Sheng-Quan, X. Diversity, abundance and distribution of grasshopper species (Orthoptera: Acrididea) in three different types of vegetation with different levels of anthropogenic disturbances in the Littoral Region of Cameroon, Journal of Insect Biodiversity. 2020, 14(1), 16-33. |
| [141] | Béguinot, J. Analysing the Role of Environmental Stresses on Species Richness and the Process of Hierarchical Structuring of Species Abundances in Marine Gastropods communities at Suva (Fiji Islands), International Journal of Environment and Climate Change Research. 2018, 8(3), 200-233. |
| [142] | Daget, J. Les modèles mathématiques en écologie. Paris France: Masson; 1976, pp. 1-172. |
APA Style
Mbianda, A. P., Mohammadou, M., Taïmanga, Kenne, A. S., Tsekane, S. J., et al. (2025). Diversity, Abundance and the Community Structure of the Flower-Visiting Insects on Sesamum indicum L. (1753) (Scrophulariales: Pedaliaceae) in Bilone (Obala-Cameroon). American Journal of Entomology, 9(1), 28-54. https://doi.org/10.11648/j.aje.20250901.14
ACS Style
Mbianda, A. P.; Mohammadou, M.; Taïmanga; Kenne, A. S.; Tsekane, S. J., et al. Diversity, Abundance and the Community Structure of the Flower-Visiting Insects on Sesamum indicum L. (1753) (Scrophulariales: Pedaliaceae) in Bilone (Obala-Cameroon). Am. J. Entomol. 2025, 9(1), 28-54. doi: 10.11648/j.aje.20250901.14
AMA Style
Mbianda AP, Mohammadou M, Taïmanga, Kenne AS, Tsekane SJ, et al. Diversity, Abundance and the Community Structure of the Flower-Visiting Insects on Sesamum indicum L. (1753) (Scrophulariales: Pedaliaceae) in Bilone (Obala-Cameroon). Am J Entomol. 2025;9(1):28-54. doi: 10.11648/j.aje.20250901.14
@article{10.11648/j.aje.20250901.14,
author = {Auguste Pharaon Mbianda and Moukhtar Mohammadou and Taïmanga and Andrea Sarah Kenne and Sedrick Junior Tsekane and Alice Virginie Tchiaze Ifoue and Xavier Arthur Nyoumi Ongolo and Dounia Dounia and Nadine Esther Otiobo Atibita and Chantal Douka and Joseph Blaise Pando and Fernand-Nestor Tchuenguem Fohouo and Martin Kenne},
title = {Diversity, Abundance and the Community Structure of the Flower-Visiting Insects on Sesamum indicum L. (1753) (Scrophulariales: Pedaliaceae) in Bilone (Obala-Cameroon)
},
journal = {American Journal of Entomology},
volume = {9},
number = {1},
pages = {28-54},
doi = {10.11648/j.aje.20250901.14},
url = {https://doi.org/10.11648/j.aje.20250901.14},
eprint = {https://article.sciencepublishinggroup.com/pdf/10.11648.j.aje.20250901.14},
abstract = {In order to identify flower-visiting insects on sesame plants and characterize the community structure, ecological survey was conducted in Bilone agroecological farm in 2022 and 2023, in 15 experimental plots (6x5.5 m each) each year, created in a 1,600 m² area. Insects were captured, stored in papillotes (Lepidoptera) or in vials containing 70° alcohol (other adults) and identified at the species level in laboratory. A total of 1,703 specimens were captured. They belonged to five orders, 12 families, 18 genera and 19 species. Hymenoptera was mostly collected order (91.5%) followed by Diptera (4.5%), Lepidoptera (1.8%), Neuroptera (0.9%) and Orthoptera (1.3%). Apidae was the most collected family (42.4%) followed by Formicidae (34.1%), Megachilidae (11.6%) while other families were rare: Acrididae (1.3%), Ascalapidae (0.9%), Calliphoridae (0.5%), Eumenidae (0.7%), Halictidae (2.2%), Muscidae (4.0%), Nymphalidae (1.3%), Pieridae (0.5%), and Vespidae (0.6%). Apis mellifera adansonii (Apidae: 30.6%) was the most recorded species, followed by Paratrechina longicornis (Formicidae: 12.3%), Pheidole megacephala (Formicidae: 9.4%), Myrmicanioa opaciventris (Formicidae: 8.9%), Megachile cincta (Megachilidae: 7.0%), Amegilla calens (Apidae: 6.2%), Xylocopa olivacea (Apidae: 5.6%), Megachile kamerunensis (Megachilidae: 4.6%), Musca domestica (Diptera: 4.0%), Camponotus maculatus (Formicidae: 3.65%), Lasioglossum hancocki (Halictidae: 2.2%), and Pteropera carnapi (Acrididae: 1.3%). Calliphora vicina (Calliphoridae) was recorded exclusively in 2022. Two exotic Diptera (Cl. vicina and Mu. domerstica) were myiasigenic species. The exotic Eumenidae Delta sp. and the afrotropical predator Ascalaphus africanus (Ascalapidae) were recorded as well as the phytophagous Acrididae Pe. carnapi. Potential pests (Nymphalidae, Pieridae and Acrididae) cumulatively represented 3.1% of the collection. The community was highly diversed and lowly dominated by a few species. Ca. maculatus was simply abundant in 2023. Amegilla calens, Ap. mellifera adansonii, Me. cincta, Me. kamerunensis, Mu. domestica, My. opaciventris, Pa. longicornis, Ph. megacephala and Xy. olivacea were simply abundant. Amegilla calens and Me. cincta, were co-dominants in 2022. Ca. maculatus and Me. kamerunensis were co-dominants in 2023. Apis mellifera adansonii, Pa. longicornis, Ph. megacephala, My. opaciventris and Xy. olivacea were co-dominants in each year. Ca. maculates and Cl. vicina were rare in 2022. Bicyclus dorothea (Nymphalidae), Delta sp. and La. hancocki were rare in 2023. Acraea acerata (Nymphalidae), Ascalaphus africanus (Ascalapidae), Catopsilia florella (Pieridae), Pteropera carnapi (Acrididae) and Synagris conuta (Vespidae) were rare. High value of Motomura constant (m=0.777 in 2022) and Preston constant (m=0.726 in 2023) suggested least evolved pioneer assemblages with species competition limited to the physical space. Overall, flower visiting insects exhibited a global positive net association.
},
year = {2025}
}
TY - JOUR T1 - Diversity, Abundance and the Community Structure of the Flower-Visiting Insects on Sesamum indicum L. (1753) (Scrophulariales: Pedaliaceae) in Bilone (Obala-Cameroon) AU - Auguste Pharaon Mbianda AU - Moukhtar Mohammadou AU - Taïmanga AU - Andrea Sarah Kenne AU - Sedrick Junior Tsekane AU - Alice Virginie Tchiaze Ifoue AU - Xavier Arthur Nyoumi Ongolo AU - Dounia Dounia AU - Nadine Esther Otiobo Atibita AU - Chantal Douka AU - Joseph Blaise Pando AU - Fernand-Nestor Tchuenguem Fohouo AU - Martin Kenne Y1 - 2025/01/21 PY - 2025 N1 - https://doi.org/10.11648/j.aje.20250901.14 DO - 10.11648/j.aje.20250901.14 T2 - American Journal of Entomology JF - American Journal of Entomology JO - American Journal of Entomology SP - 28 EP - 54 PB - Science Publishing Group SN - 2640-0537 UR - https://doi.org/10.11648/j.aje.20250901.14 AB - In order to identify flower-visiting insects on sesame plants and characterize the community structure, ecological survey was conducted in Bilone agroecological farm in 2022 and 2023, in 15 experimental plots (6x5.5 m each) each year, created in a 1,600 m² area. Insects were captured, stored in papillotes (Lepidoptera) or in vials containing 70° alcohol (other adults) and identified at the species level in laboratory. A total of 1,703 specimens were captured. They belonged to five orders, 12 families, 18 genera and 19 species. Hymenoptera was mostly collected order (91.5%) followed by Diptera (4.5%), Lepidoptera (1.8%), Neuroptera (0.9%) and Orthoptera (1.3%). Apidae was the most collected family (42.4%) followed by Formicidae (34.1%), Megachilidae (11.6%) while other families were rare: Acrididae (1.3%), Ascalapidae (0.9%), Calliphoridae (0.5%), Eumenidae (0.7%), Halictidae (2.2%), Muscidae (4.0%), Nymphalidae (1.3%), Pieridae (0.5%), and Vespidae (0.6%). Apis mellifera adansonii (Apidae: 30.6%) was the most recorded species, followed by Paratrechina longicornis (Formicidae: 12.3%), Pheidole megacephala (Formicidae: 9.4%), Myrmicanioa opaciventris (Formicidae: 8.9%), Megachile cincta (Megachilidae: 7.0%), Amegilla calens (Apidae: 6.2%), Xylocopa olivacea (Apidae: 5.6%), Megachile kamerunensis (Megachilidae: 4.6%), Musca domestica (Diptera: 4.0%), Camponotus maculatus (Formicidae: 3.65%), Lasioglossum hancocki (Halictidae: 2.2%), and Pteropera carnapi (Acrididae: 1.3%). Calliphora vicina (Calliphoridae) was recorded exclusively in 2022. Two exotic Diptera (Cl. vicina and Mu. domerstica) were myiasigenic species. The exotic Eumenidae Delta sp. and the afrotropical predator Ascalaphus africanus (Ascalapidae) were recorded as well as the phytophagous Acrididae Pe. carnapi. Potential pests (Nymphalidae, Pieridae and Acrididae) cumulatively represented 3.1% of the collection. The community was highly diversed and lowly dominated by a few species. Ca. maculatus was simply abundant in 2023. Amegilla calens, Ap. mellifera adansonii, Me. cincta, Me. kamerunensis, Mu. domestica, My. opaciventris, Pa. longicornis, Ph. megacephala and Xy. olivacea were simply abundant. Amegilla calens and Me. cincta, were co-dominants in 2022. Ca. maculatus and Me. kamerunensis were co-dominants in 2023. Apis mellifera adansonii, Pa. longicornis, Ph. megacephala, My. opaciventris and Xy. olivacea were co-dominants in each year. Ca. maculates and Cl. vicina were rare in 2022. Bicyclus dorothea (Nymphalidae), Delta sp. and La. hancocki were rare in 2023. Acraea acerata (Nymphalidae), Ascalaphus africanus (Ascalapidae), Catopsilia florella (Pieridae), Pteropera carnapi (Acrididae) and Synagris conuta (Vespidae) were rare. High value of Motomura constant (m=0.777 in 2022) and Preston constant (m=0.726 in 2023) suggested least evolved pioneer assemblages with species competition limited to the physical space. Overall, flower visiting insects exhibited a global positive net association. VL - 9 IS - 1 ER -
Department of Plant Biology, University of Douala, Douala, Cameroon
Research Fields: Applied entomology, Insects biology, biostatistics and pollinators, Apiculture, Insects-plants interactions, Animal Ethology, Animal Ecology
Department of Biology and Physiology of Animal Organisms, University of Douala, Douala, Cameroon
Research Fields: Applied entomology, Insects biology, biostatistics and pollinators, Apiculture, Insects-plants interactions, Animal Ethology, Animal Ecology
Department of Agronomy, University of Douala, Douala, Cameroon
Research Fields: Botanical insecticides, Bees, Biological agriculture, Yield agricultural products, Cowpea production, Pollinization
Department of Biology and Physiology of Animal Organisms, University of Douala, Douala, Cameroon
Research Fields: Water toxicology, Waterborne diseases, Water quality of life, Biostatistics, Animal Ethology, Animal Ecology
Department of Biology and Physiology of Animal Organisms, University of Douala, Douala, Cameroon
Research Fields: Applied zoology, Quality of life and biostatistics, Wildlife Protection, Control of protected areas, Animal Ethology, Animal Ecology
Department of Plant Biology, University of Douala, Douala, Cameroon
Research Fields: Applied entomology, Insects biology, biostatistics, pollinators, Plant physiology, Soil fertilization
Department of Agronomy, University of Dschang, Obala, Cameroon
Research Fields: Applied entomology, Insects biology, Pollinators, Seeds production, Agriculture, Plant variety amelioration.
Higher Teacher Training College, University of Yaounde 1, Yaounde, Cameroun
Research Fields: Applied entomology, Insects biology, biostatistics and pollinators, Apiculture, Insects-plants interactions, Animal Ethology, Animal Ecology
Department of Biology and Physiology of Animal Organisms, University of Douala, Douala, Cameroon
Research Fields: Applied entomology, Insects biology, biostatistics and pollinators, Apiculture, Insects-plants interactions, Animal Ethology, Animal Ecology
Higher Teacher Training College, University of Yaounde 1, Yaounde, Cameroun
Research Fields: Applied entomology, Insects biology, biostatistics and pollinators, Apiculture, Insects-plants interactions, Animal Ethology, Animal Ecology
Higher Teacher Training College, University of Maroua, Maroua, Cameroun
Research Fields: Applied entomology, Insects biology, biostatistics and pollinators, Apiculture, Insects-plants interactions, Animal Ethology, Animal Ecology
Department of Biological Sciences, University of Ngaoundere, Ngaoundere, Cameroon
Research Fields: Applied entomology, Insects biology, biostatistics and pollinators, Apiculture, Insects-plants interactions, Animal Ethology, Animal Ecology
Department of Biology and Physiology of Animal Organisms, University of Douala, Douala, Cameroon
Research Fields: Biostatistics and Biology of the Animal Populations, Entomology and Myrmecology, Animal Ethology, Animal Ecology and sociobiology, Applied entomology and plant protection, Biological control of pest insects
Figure 1. Localization map of the study site. A: Centre Region in Cameroon [44]; B: Lekie department in the Centre Region [44]; C: Obala in the lekie department [44]; D: Distance from Obala to the Bilone agroecological farm; E: study site at Bilone agroecological farm (Google Earth Pro for windows version 7.3.4.8642).
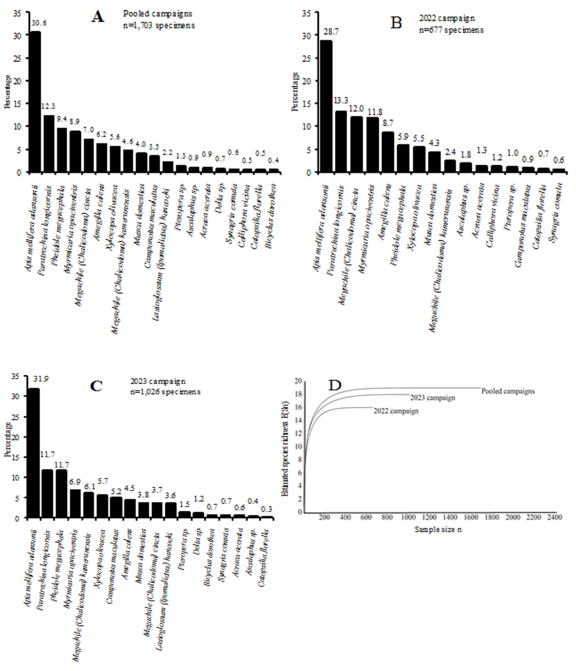
Figure 2. Rank-frequency diagrams of the collected insects in the pooled campaigns (A), in 2022 (B) and 2023 (C) showing species in order of numerical dominance. The species rarefaction curves (D) (estimated species richness as a function of the sample size variation) showed the low species richness variation in 2022 and the high species richness in the pooled campaigns.
Information

