2. Literature Review
2.1. Origin of its Name and Nature of Striga
There are a number of possible origins of the name
Striga. It could have been derived from Latin words meaning variously “strait, horshy”, lean or witch'.
Striga often refer to the word “witch”, presumably because plants infested by the parasite display stunted growth and an overall drought-like phenotype long before
Striga plants appear
| [1] | Spallek, T., Mutuku, J. M. & Shirasu, K. (2013). The genus Striga: a witch profile. Molecular Plant Pathology, 14, 861-869. |
[1]
. Parasitic plants are found in about 17 families. However, only five of them include agricultural pests
| [2] | Parker, C. (1991). Protection of crops against parasitic weeds. Crop Protection, 10: 6-22. |
[2]
. In among these families, Orobanchaceae received considerable attention, because of its relevance in world agriculture. This family is of interest in evolutionary studies, and because it encompasses closely related parasites with vast differences in their host requirements. Among all flowering plant families only Orobanchaceae in comprised of various genera ranging from completely eutrophic to specialized obligate parasites. The genus
Striga (witchweed) belongs to the Orobanchaceae family (ex Scrophulariaceae) and is obligate root hemiparasitic plants
| [4] | Yoshida, S. (2019). Genetic basis for host and parasitic plant communication. In the Proceeding of the 15th World Congress on Parasitic Plants. June 30 – July 5 2019, Amsterdam, the Netherlands. |
[4]
. This genus currently comprises of 42 species worldwide, which are parasitic by nature, of which at least 11 are known to attack crops. In compensation for its rudimentary root system,
Striga penetrates the roots of other plants and diverts essential nutrients for its growth and development
| [1] | Spallek, T., Mutuku, J. M. & Shirasu, K. (2013). The genus Striga: a witch profile. Molecular Plant Pathology, 14, 861-869. |
[1]
.
Striga plants are herbaceous. The genus is characterized by opposite leaves, irregular bright colored flowers with corolla divided into a tube spreading lobes, herbaceous habitat, small seeds, and parasitism.
S. hermonthica has bright to dark green leaves, erect and usually branched stems grow up to 77 cm or more. Stems are stout and quadrangular. Leaves are linear, lanceolate or lanceolate with actuate or acuminate tips, 1-3 in. long, very scabrous. The inflorescence possesses 6-10 open flowers that are 1-2 cm across. The flowers are pink, red, white, purple or yellow. The spike has occasionally more than 10 open flowers and the corolla normally drops a few days after fertilization
| [5] | Parker, C. & Riches, C. (1993). Parasitic Weeds of the World: Biology and Control. Wallingford CAB international. 1993 pp. 4-332. |
[5]
. The number of capsules per plant may be on the average 42-110. The number of seeds per capsule varies from about 700 in
S. hermonthica to 800 in
S. asiatica. In a single growing season, each
Striga plant is capable of producing up to 76628-minute dust like seeds which are easily dispersible by wind, crop seeds, water, people and may stay in the soil for up to 20 years
| [5] | Parker, C. & Riches, C. (1993). Parasitic Weeds of the World: Biology and Control. Wallingford CAB international. 1993 pp. 4-332. |
| [6] | Dafaallah, A. B., Babiker, A. G. T & Zain El abdeen, M. H. (2015). Variability in Striga hermonthica (Del.) Benth, Populations in Gadarif Area, Eastern Sudan. Sudan University of Science and Technology Journal of Agricultural and Veterinary Sciences, 16(2), 119-132. |
[5, 6]
. Seeds of
S. hermonthica are extremely small, about 0.15 x 0.31 mm, 0.38 mm diameter and weighing about 3-15 µg.
2.2. Distribution of Striga
The economically important root-parasitic weeds have their center of origin in the old world. Africa was described as the place of origin of the agriculturally important genera of the family Orobanchaceae
| [8] | Ejeta, G. H. (2007). Breeding for Striga Resistance in Sorghum: Exploitation of an Intricate Host–Parasite Biology. Crop science 47(S3), S216–S227. |
[8]
. The species are found in most regions south of the Sahara except in areas, where precipitation is too high or temperatures are too low for development
| [9] | Ejeta, G. & Gressel, J. (2007) Integrating New Technologies for Striga Control: Towards Ending the Witch-Hunt. Singapore; Hackensack, NJ: World Scientific. |
[9]
. The parasite prevails from sea-level up to 2,000 meters above sea level and in almost all soil types
| [7] | Oswald, A. (2005) Striga control - technologies and their dissemination. Crop Protection, 24. 333-342. |
[7]
.
Striga was thought to have originated in the vast tropical areas of the savannah between the Simien Mountains of Ethiopia and the Nuba hills in Sudan
| [10] | Atera, E. & Itoh, K. (2011) Evaluation of ecologies and severity of Striga weed on rice in sub-Saharan Africa. Agric. Biol. J. N. Am. 2, 752–760. |
[10]
. This region has also been reported to be the center of origin of domesticated sorghum (
Sorghum bicolor L.).
Striga species are sensitive to water logging and are more prevalent in poor soils than on fertile soils. Surveys conducted in 2007 indicated that the parasite is spreading rapidly
| [15] | Babiker, A. G. T. (2007). Striga: The Spreading Scourge in Africa. Regulation of Plant Growth and development, 42, 74-87. |
[15]
. Economically important
Striga species have broad distribution setting conditions for genetically structured populations based on geographic locations
| [13] | Mohamed, K., Bolin, J., Musselman, L. & Peterson, A. (2007). Genetic diversity of Striga and implications for control and modeling future distributions. In: Ejeta, G. Gressel, J. (eds.) Integrating New Technologies for Striga Control: Towards Ending the Witch-hunt. 71-84. |
[13]
.
Striga hermonthica is mainly distributed throughout the semi-arid area of northern tropical Africa, but extends into south west Arabia and south tropical Africa; including Angola, Namibia and Malagasy. It is less widespread than
S. asiatica in southern tropical Africa
| [5] | Parker, C. & Riches, C. (1993). Parasitic Weeds of the World: Biology and Control. Wallingford CAB international. 1993 pp. 4-332. |
[5]
.
2.3. Physiology of Striga
Many aspects of Striga biology are not fully understood. They include the physiological processes of photosynthesis, respiration, transpiration, water relations, and cause of heavy crop-yield reductions, morphology, and analoging of the haustorium in relation to its function
| [25] | Nasreldin Mohemed Ahmed (2018): The role of strigolactones in resistance, tolerance and control of Striga infection in Sorghum; PhD Thesis Dissertation submitted in fulfillment of the requirements for the degree of doctor at Wageningen University. |
[25]
. The rates of photosynthesis of Striga species were found to be very low in contrast to the respiration and transpiration rates of higher plants. Stomata of drought-stressed Striga plants were found to be open in darkness and their control is poor. This high rate of transpiration is interpreted as a means of maximizing solute transfer from host to parasite. Such reliance on high rates of transpiration has suggested use of anti-transpirants in Striga control
| [27] | Haussmann, B. I. G., Hess, D. E. Weiz, H. G. and Geiger, H. H. 2000. Improved methodologies for Breeding, Striga resistant sorghums, Field Crops Research. |
[27]
.
2.4. Host Range of Striga Species and Its Life Cycle
Striga hermonthica is the most widespread and most damaging among
Striga species. The range of crops attacked by
S. hermonthica includes all major tropical cereals. Occurrence is also reported on some temperate crops, including teff (
Eragrotis tef (Zuccagni) Trotter) and barley (
Hordeum vulgare L.)
| [5] | Parker, C. & Riches, C. (1993). Parasitic Weeds of the World: Biology and Control. Wallingford CAB international. 1993 pp. 4-332. |
[5]
. The main cereal crops parasitized by
S. hermonthica are sorghum, millet, maize, rice and sugarcane (
Saccharum officinarum L.)
.
Striga aspera has been recorded on most of the major cereal crops,
i.e. maize, sorghum and sugarcane. Host crops attacked by
S. asiatica are virtually the same as those for
S. hermonthica, sorghum and maize being the most widely
damaged especially in India and Southern Africa
| [5] | Parker, C. & Riches, C. (1993). Parasitic Weeds of the World: Biology and Control. Wallingford CAB international. 1993 pp. 4-332. |
[5]
.
S. gesnerioides has an extensive host range that includes
species of the family
Fabaceae, Acanthaceae,
Convolvulaceace, Euphorbiaceae,
Redaliaceae and
Solanaceae. However, this parasitic weed is only of economic significance in cowpea (
Vigna unguiculata (L.) Walpe.). In West Africa and revealed the sporadic occurrence on tobacco (
Nicotiana tabacum L.) in South Africa and rarely on sweet potato (
Ipomoea batatas (L.) Lam.) in south Africa
| [11] | Parker. C. (2012). Parasitic weeds: A world challenge. Weed Sci, 60. 269–276. |
[11]
.
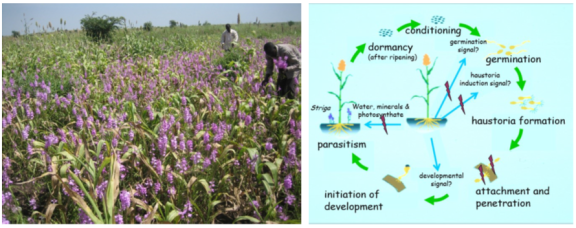
Figure 1. The Striga life cycle showing intricate association between the parasite, its hosts, and the environment.
Striga’s complete dependence upon a host for survival requires close coordination of its life cycle with that of the host.
Striga seeds are minute with limited stored food reserves and
Striga germilings survive only for about three days unless attachment to a host root is achieved
| [8] | Ejeta, G. H. (2007). Breeding for Striga Resistance in Sorghum: Exploitation of an Intricate Host–Parasite Biology. Crop science 47(S3), S216–S227. |
[8]
. It is not surprising, therefore, that germination of
Striga seeds is under control of the host through chemical signals exuded from its roots, so that germination usually occurs only when a plant root is available. Subsequent to germination, haustorium initiation occurs in response to a second host-derived signal
| [25] | Nasreldin Mohemed Ahmed (2018): The role of strigolactones in resistance, tolerance and control of Striga infection in Sorghum; PhD Thesis Dissertation submitted in fulfillment of the requirements for the degree of doctor at Wageningen University. |
[25]
. The haustorium penetrates the cortex, guided possibly by the internal chemistry of the host root and establishes connection with the vascular system. Following connection with the host xylem, the plumular end comes out of the seed coat and further development occurs. The life cycle of the parasite is divided into a non-parasitic or vegetative phase and a parasitic mode. The non-parasitic mode includes the processes of after-ripening, conditioning and germination. The parasitic mode starts with the initiation of a haustorium from the vegetative to the parasitic mode of life
| [17] | Yoneyama, K., Awad, A. A., Xie, X., Yoneyama, K. & Takeuchi, Y. (2010). Strigolactones as germination stimulants for root parasitic plants. Plant Cell Physiol. 51, 1095-1103. |
[17]
.
Non-parasitic Mode (Vegetative Phase)
After- ripening: The seeds of parasitic weeds are tiny relative to those of free- living angiosperms. Energy reserves in small seeds are limited and sufficient for a short period of autonomous growth.
Striga seeds have an after-ripening requirement and cannot germinate in the season in which they were produced
| [18] | Riches, C. R. & Ejeta, G. (2007). Biology of host – parasite interaction in Striga species. In: Ejeta, G. and Gressel, J. (eds.) Integrating New Technologies for Striga Control: Towards Ending the Witch-hunt. pp. 19-32. |
[18]
. This requirement is an evolutionary adaptation to prevent newly matured
Striga seeds from germinating too late in a growing season, when host plants are normally senescing and are not capable of supporting a parasitic plant to maturity
| [28] | Temesgen Teressa, (2019). “Review on Striga Distribution, Infestation and Genetic Potential in Ethiopian Sorghum (Sorghum Bicolor (L.) Moench)” International Journal of Research Studies in Agricultural Sciences (IJRSAS), 5(2), pp. 23-31. |
[28]
. A difference in the length of after-ripening periods exists between
Striga species. The after-ripening process is described as a means of adaptation of
Striga to the semi-arid climate
| [22] | Doggett, H. (1988). Sorghum. (second edition) John-Wiley and Sons, New York, London, pp. 512. |
[22]
. Warm and dry conditions are pre-requisites for after ripening
| [23] | Babiker, A. G. T, Ma Y, Sugimoto, Y. & Inanaga, S. (2000) Conditioning period, Co2 and GR24 influence ethylene biosynthesis and germination of Striga hermonthica. Plant Physiology, 109, 75–80. |
[23]
.
Conditioning: After-ripened, seeds will not germinate until they have passed through a preconditioning period. A complication in the germination of
S. hermonthica and other
Striga species is their inability to germinate, even in the presence of a suitable stimulant, until they receive a pre-treatment period in warm and moist conditions (conditioning or preconditioning) for at least a few days, ideally 1-2 weeks. The optimum temperature for conditioning is between 25 and 35
0C for
S. hermonthica | [16] | Berhane Sibhatu Review on Striga Weed Management, 2016 Department of Agronomy, Ethiopian Institute of Agricultural Research, Mehoni Agricultural Research Center. |
[16]
. The duration and temperature optimum for the conditioning period of
Striga seeds vary with species. In
S. asiatica the optimum conditioning period is 21 days at 22 ºC and is two weeks at 33 ºC for
S. hermonthica. Higher temperatures will, however, result in rapid conditioning of this species, but percentage germination will not be as high even after several weeks
| [9] | Ejeta, G. & Gressel, J. (2007) Integrating New Technologies for Striga Control: Towards Ending the Witch-Hunt. Singapore; Hackensack, NJ: World Scientific. |
[9]
.
Seed germination:
Striga seeds germinate only when they receive an exogenous stimulant subsequent to conditioning. The natural stimulant is exuded by the host’s roots and some non-host plants. Following germination, the radicles grow towards the host roots, indicating a chemotrophic effect
| [25] | Nasreldin Mohemed Ahmed (2018): The role of strigolactones in resistance, tolerance and control of Striga infection in Sorghum; PhD Thesis Dissertation submitted in fulfillment of the requirements for the degree of doctor at Wageningen University. |
[25]
. Several germination stimulants have been isolated and include strigolactones, dihydrosorogoleone, sesquiterpene, kinetin, coumarin, jasmonate, ethylene and fungal metabolites. A large number of investigators have attempted to isolate, characterize and/ or identify the stimulant from many hosts and non-host plants
| [7] | Oswald, A. (2005) Striga control - technologies and their dissemination. Crop Protection, 24. 333-342. |
[7]
. The natural stimulants are highly active, but are present in root exudates in such extremely low levels that their isolation, purification and identification have been difficult
| [14] | Musselman, L. J. & Hepper, F. (1986). The witch-weed (Striga, Schrophulariaceae) of the Sudan Republic. Kew Bullettin, 41, 205-221. |
[14]
. The first natural germination stimulant is “Strigol”. Strigol was isolated from cotton [
Gossypium hirsutum (L.)], non-host plant. Strigol is active on the
S. asiatica at 10
-16 M. Several years after the discovery of strigol several natural germination stimulants were identified from the roots of sorghum
| [27] | Haussmann, B. I. G., Hess, D. E. Weiz, H. G. and Geiger, H. H. 2000. Improved methodologies for Breeding, Striga resistant sorghums, Field Crops Research. |
[27]
. Five different stimulants, strigol, strigyl acetate, sorgolactone, alectrol and orobanchol were isolated from host and non- host plants. These compounds, because of similarity in chemical structure, are collectively referred as to strigolactones (
Figure 1). Strigolactones are associated with the negative regulation of root and shoot branching (tillering). They also induce hyphal branching of arbuscular mycorrhizal (AM) fungi, presumably to attract them in lownutrient environments. It is at least known that most of the important
Striga species will respond to strigol and to the “Strigol analogues” that have been synthesized and tested as possible means of control
| [17] | Yoneyama, K., Awad, A. A., Xie, X., Yoneyama, K. & Takeuchi, Y. (2010). Strigolactones as germination stimulants for root parasitic plants. Plant Cell Physiol. 51, 1095-1103. |
[17]
. Genes encoding the key enzymes in ethylene biosynthesis, ACC synthase and ACC oxidase are regulated by germination stimulants and conditioning
| [28] | Temesgen Teressa, (2019). “Review on Striga Distribution, Infestation and Genetic Potential in Ethiopian Sorghum (Sorghum Bicolor (L.) Moench)” International Journal of Research Studies in Agricultural Sciences (IJRSAS), 5(2), pp. 23-31. |
[28]
.
2.5. The Parasitic Mode: Contact and Attachment (Haustorial Initiation)
Once the seeds of
Striga have germinated up to several mms (2-4 mm) from a host root, radicle has to come in contact with the host root in order to parasitize it. Given the purely random directional growth after germination, it has been estimated that the chance of contact with a single root with small diameter may be less than 10%
| [8] | Ejeta, G. H. (2007). Breeding for Striga Resistance in Sorghum: Exploitation of an Intricate Host–Parasite Biology. Crop science 47(S3), S216–S227. |
| [9] | Ejeta, G. & Gressel, J. (2007) Integrating New Technologies for Striga Control: Towards Ending the Witch-Hunt. Singapore; Hackensack, NJ: World Scientific. |
[8, 9]
. It is to be expected that in the course of evolution these chances might have been improved by the development of some chemotropic or other direction-seeking assistance. Indeed, reference
| [25] | Nasreldin Mohemed Ahmed (2018): The role of strigolactones in resistance, tolerance and control of Striga infection in Sorghum; PhD Thesis Dissertation submitted in fulfillment of the requirements for the degree of doctor at Wageningen University. |
[25]
presented evidence for chemotropism in
S. hermonthica, while
| [20] | Saunders, A. (1933). Studies in phenorogamic parasitism with particular reference to Striga lutea (Lour). South Africa Department of Agriculture, Science Bulletin, 128, 1-57. |
[20]
observed the phenomenon in
S. asiatica. Effective chemotropism could result from a gradient in pH around the root or an inhibitory effect of the root exudates on the side of the radicle nearest to host root. Witch weed germilings survive only for about three to seven days unless attachment to a host root is achieved
| [3] | Babiker, A. G. T, Ejeta, G., Butler, L. G. & Woodson, W. R. (1993). Ethylene biosynthesis and strigol-induced germination of Striga asiatica. Physiologia Plantarium, 88,359-365. |
[3]
.
The haustorium penetrates the host root, establishes connection with host xylem, guided possibly by host derived secondary metabolites. Unlike its response to germination stimulants and haustorium initiators,
Striga is non-specific with response to the attachment. Attachment frequencies were reported to be similar for host and nonhost plant species
| [18] | Riches, C. R. & Ejeta, G. (2007). Biology of host – parasite interaction in Striga species. In: Ejeta, G. and Gressel, J. (eds.) Integrating New Technologies for Striga Control: Towards Ending the Witch-hunt. pp. 19-32. |
[18]
. The process of haustorial development and penetration of the host is similar in
S. hermonthica | [27] | Haussmann, B. I. G., Hess, D. E. Weiz, H. G. and Geiger, H. H. 2000. Improved methodologies for Breeding, Striga resistant sorghums, Field Crops Research. |
[27]
and
S. asiatica Sticky hairs on the young haustorium help the parasite germiling to adhere to any surface. After attachment by these hairs, intrusive cells develop at the root tip and penetrate the cortex of the host.
2.5.1. Penetration and Establishment of the Parasite
Penetration is aided by enzymatic secretion leading to separation of the host cortex cells. The haustorium sometimes fails to complete its penetration of the cortex and may also fail to cross the endodermis which sometimes provides a barrier. In
S. asiatica, the time from the first penetration of the epidermis to an established connection with the host stele is 60 hours
| [28] | Temesgen Teressa, (2019). “Review on Striga Distribution, Infestation and Genetic Potential in Ethiopian Sorghum (Sorghum Bicolor (L.) Moench)” International Journal of Research Studies in Agricultural Sciences (IJRSAS), 5(2), pp. 23-31. |
[28]
. Following a successful connection to the host xylem, the plumular end of the seedling emerges from the seed coat and
Striga becomes fully dependent on the host. The successful parasite establishment creates a strong nutrients sink leading, to drastic reductions in host growth and yield
| [20] | Saunders, A. (1933). Studies in phenorogamic parasitism with particular reference to Striga lutea (Lour). South Africa Department of Agriculture, Science Bulletin, 128, 1-57. |
[20]
. On emerging from the soil, the aerial parts of the parasite turn green
Striga plants begin to photosynthesize. However, the low CO2 fixation and high dark respiration rates of
Striga result in a negative carbon gain over the 24-h period, thus making the parasite still unable to survive in the absence of host attachment.
Striga is described by the high transpiration rates. These rates suggest that most host photo assimilates are obtained by transpirational pull, explaining why high humidity is inhibitory to
Striga growth. Indeed,
Striga stomata show high conductance and respiration rates and little response to dark-induced closure
| [27] | Haussmann, B. I. G., Hess, D. E. Weiz, H. G. and Geiger, H. H. 2000. Improved methodologies for Breeding, Striga resistant sorghums, Field Crops Research. |
[27]
.
Flowering time is species and environment dependent.
S. gesnerioides begins to flower as it emerges.
S. hermonthica,
S. asiatica and
S. aspera begin to flower 4 weeks after
emergence. Flowering begins basally on the raceme, and
seeds are mature 4 weeks after flowering
| [5] | Parker, C. & Riches, C. (1993). Parasitic Weeds of the World: Biology and Control. Wallingford CAB international. 1993 pp. 4-332. |
[5]
.
When conditions are favorable for parasite growth, the
parasite will normally germinate and attach to the root
system within 2-3 weeks after host germination, emerge
after 4-7 weeks and flower within 7-8 weeks. Viable seeds
are probably produced within 2 weeks of flower opening
and are fully matured and shed about 2 weeks later
| [22] | Doggett, H. (1988). Sorghum. (second edition) John-Wiley and Sons, New York, London, pp. 512. |
[22]
. The
minimal length of the life cycle of the parasite, from
germination to seed production comprises an average of 4
months
| [15] | Babiker, A. G. T. (2007). Striga: The Spreading Scourge in Africa. Regulation of Plant Growth and development, 42, 74-87. |
[15]
.
2.5.2. Striga Species Exhibit Variation in their Mode of Reproduction
S. hermonthica and
S. gesneriodes are allogamous that is they observe cross pollination and usually rely on vectors such as bees and other agents of pollination for pollen transfer
| [24] | Dafaallah B. Awadallah (2019): Biology and Physiology of Witchweed (Striga spp.): A Review, International Journal of Academic Multidisciplinary Research, 3(10): 42-51. 25. |
| [28] | Temesgen Teressa, (2019). “Review on Striga Distribution, Infestation and Genetic Potential in Ethiopian Sorghum (Sorghum Bicolor (L.) Moench)” International Journal of Research Studies in Agricultural Sciences (IJRSAS), 5(2), pp. 23-31. |
[24, 28]
.
S. asiatica on the other hand is autogamous that is it observes self-pollination and so, no vectors are needed for pollination instead pollens are picked by the elongation of style and fertilization takes place
| [1] | Spallek, T., Mutuku, J. M. & Shirasu, K. (2013). The genus Striga: a witch profile. Molecular Plant Pathology, 14, 861-869. |
[1]
. The development of the Striga spp. is influenced by the Soil type, soil temperature, tillage systems and the parasite thrives best under conditions of mono-cropping of susceptible host
| [5] | Parker, C. & Riches, C. (1993). Parasitic Weeds of the World: Biology and Control. Wallingford CAB international. 1993 pp. 4-332. |
[5]
.
As much as half of the
Striga life cycle is subterranean, growing completely at the expense of its host and the parasite inflicts most of its damage to the host during this phase of its life cycle. Symptoms displayed by infected hosts, include stunting, toxic' effects, reduction of internode expansion, wilting, chlorosis, increased root: shoot ratio, reduced photosynthetic rate and decreased growth and yield. Parasitism by
S. hermonthica leads to perturbation of hormonal balance and a marked change in the amino acid content of the grains
| [6] | Dafaallah, A. B., Babiker, A. G. T & Zain El abdeen, M. H. (2015). Variability in Striga hermonthica (Del.) Benth, Populations in Gadarif Area, Eastern Sudan. Sudan University of Science and Technology Journal of Agricultural and Veterinary Sciences, 16(2), 119-132. |
[6]
.
3. Breeding Approaches of Sorghum to Striga Resistant
3.1. Conventional Breeding Approach
Both inter-specific variability among Striga species and intra-specific variation for aggressiveness must be taken into account when breeding for striga resistance
| [26] | Ejeta G, Butler LG, Babiker AG (1992) New approaches to the control of Striga. Striga Research at Purdue University, Research Bulletin RB-991. Agricultural Experiment Station, Purdue University, West Lafayette, IN. |
[26]
. In order to obtain stable, polygenic resistance, breeding materials should be evaluated at various locations with different striga populations or host-specific races
| [9] | Ejeta, G. & Gressel, J. (2007) Integrating New Technologies for Striga Control: Towards Ending the Witch-Hunt. Singapore; Hackensack, NJ: World Scientific. |
[9]
. In doing so, quarantine regulations must be strictly respected, and striga species or strains should not be introduced into regions where they do not already occur. If seed shortage imposes a constraint on progeny evaluation, a reduction in plot size should be preferred over reduction of the number of test locations, since there is always the danger of losing data from one location due to “non striga years'' or other obstacles. The breeder may also consider a trade-off between numbers of replications versus number of sites; however, the number of replications should not fall below four. To avoid seed shortage and therefore a trade-off between replications and sites, breeders could use inbred generations as test entries In addition to multi-location testing, the following breeding measures have been put forward by groups active in the field
| [18] | Riches, C. R. & Ejeta, G. (2007). Biology of host – parasite interaction in Striga species. In: Ejeta, G. and Gressel, J. (eds.) Integrating New Technologies for Striga Control: Towards Ending the Witch-hunt. pp. 19-32. |
| [26] | Ejeta G, Butler LG, Babiker AG (1992) New approaches to the control of Striga. Striga Research at Purdue University, Research Bulletin RB-991. Agricultural Experiment Station, Purdue University, West Lafayette, IN. |
[18, 26]
. Characterize crop germplasm, search for sources of resistance and tolerance in elite material, and improve currently available sources of resistance for agronomic performance:
1) Include wild relatives with superior resistance in the breeding program;
2) Transfer resistance genes into productive, well adapted genotypes;
3) Pyramid resistance genes to obtain more durable and stable, polygenic resistance;
4) Combine lines with different resistance mechanisms to form hybrids or synthetics, to increase durability of resistance;
5) Develop breeding populations with multiple sources of resistance using recurrent selection procedures;
6) Develop and employ marker-assisted selection techniques for broad-based, quantitative striga resistance under field conditions.
Sorghum, due to the availability of nuclear and cytoplasmic-genetic male sterility, offers a wide range of possible genetic structures to the breeder, including homozygous lines, homogeneous or heterogeneous hybrids, as well as homo or heterozygous, heterogeneous population or synthetic varieties. The potential merit of heterozygous sorghum cultivars was demonstrated by the average superiority of F2 populations over their parental lines of 18% for grain yield under striga infestation, averaged across four locations in Mali and Kenya
| [27] | Haussmann, B. I. G., Hess, D. E. Weiz, H. G. and Geiger, H. H. 2000. Improved methodologies for Breeding, Striga resistant sorghums, Field Crops Research. |
[27]
. In addition,
| [21] | Parker, C. (1984). The influence of Striga species on sorghum under varying nitrogen fertilization. In: Parker, C. Musselman, L. J., Polhill, R. M. and Wilson, A. K. (eds.) Proceedings, 3rd International Symposium on Parasitic Weeds. Aleppo, Syria, 1984. ICARDA, Aleppo. pp. 90-98. |
[21]
reported that hybrid vigor can provide a degree of tolerance to striga in sorghum and maize, which is reflected in reduced yield depression under conditions of striga infestation.
Sorghum hybrids were reported to out yield parental lines or local varieties under variable drought stress in semi-arid, striga-free areas of East and West Africa
| [20] | Saunders, A. (1933). Studies in phenorogamic parasitism with particular reference to Striga lutea (Lour). South Africa Department of Agriculture, Science Bulletin, 128, 1-57. |
[20]
. Instead of hybrids, other types of cultivars could be produced which capitalize on heterozygosity, e.g., synthetics built up from components with high out crossing rates and superior combining ability for striga resistance and grain yield. A synthetic cultivar can be re-grown for a few seasons without serious changes in its genetic composition, which is convenient for the small-scale farmers
| [28] | Temesgen Teressa, (2019). “Review on Striga Distribution, Infestation and Genetic Potential in Ethiopian Sorghum (Sorghum Bicolor (L.) Moench)” International Journal of Research Studies in Agricultural Sciences (IJRSAS), 5(2), pp. 23-31. |
[28]
. The lack of reliable single-plant screening techniques in the field generally causes selection for striga resistance to be deferred until true-breeding progenies are available. This means that large numbers of progeny have to be advanced before the trait of interest can be assessed, a time- and cost-intensive procedure.
The agar-gel assay
| [9] | Ejeta, G. & Gressel, J. (2007) Integrating New Technologies for Striga Control: Towards Ending the Witch-Hunt. Singapore; Hackensack, NJ: World Scientific. |
[9]
is an excellent tool to transfer the low stimulant character to locally adapted cultivars using classical back-cross procedures. The fact that the low stimulant gene (s) were reported to be recessive renders the back-cross program more complicated and time-consuming. With its high heritability and the possibility to screen large numbers of entries, the
in vitro germination distance fulfills two major prerequisites for an indirect selection trait. Coefficients of correlation between germination distance and striga resistance under field conditions are generally positive but vary among genetic materials and test locations
| [5] | Parker, C. & Riches, C. (1993). Parasitic Weeds of the World: Biology and Control. Wallingford CAB international. 1993 pp. 4-332. |
| [8] | Ejeta, G. H. (2007). Breeding for Striga Resistance in Sorghum: Exploitation of an Intricate Host–Parasite Biology. Crop science 47(S3), S216–S227. |
[5, 8]
Breeders should bear in mind that screening for individual resistance mechanisms in the laboratory could result in a loss of valuable materials possessing resistance mechanisms other than those evaluated. The risk increases with increasing selection intensity, i.e., with a reduced effective population size. One strategy could be to use laboratory assays for individual resistance mechanisms as an initial screening of a larger number of breeding materials, followed by the more resource-demanding field screening. This would offer the possibility to identify resistance sources with multiple resistance mechanisms. Networking and exchange of useful materials are also important steps towards more efficient breeding programs for resistance to striga in sorghum.
3.2. Marker Assisted Breeding; Molecular Marker Techniques Are a Powerful New Tool in Plant Breeding
They permit identification and mapping of genes for individual, monogenic resistance mechanisms (like the low stimulant locus) and of quantitative trait loci (QTL) involved in polygenic, quantitative resistance under field conditions. The utility of DNA markers in resistance breeding depends on the existence of tight linkage between these markers and the resistance genes or QTL of interest. In marker-assisted breeding programs, such linkage allows the breeder to select for resistance by identifying the DNA marker instead of evaluating the materials directly for resistance traits
| [2] | Parker, C. (1991). Protection of crops against parasitic weeds. Crop Protection, 10: 6-22. |
[2]
. The integration of molecular marker selection techniques into plant breeding promises a more rapid incorporation of desirable genes into improved cultivars, and facilitates the transfer of novel genes from related wild species
| [9] | Ejeta, G. & Gressel, J. (2007) Integrating New Technologies for Striga Control: Towards Ending the Witch-Hunt. Singapore; Hackensack, NJ: World Scientific. |
| [25] | Nasreldin Mohemed Ahmed (2018): The role of strigolactones in resistance, tolerance and control of Striga infection in Sorghum; PhD Thesis Dissertation submitted in fulfillment of the requirements for the degree of doctor at Wageningen University. |
[9, 25]
. Detecting resistance genes by their linkage to DNA markers makes it possible to screen for many different resistance genes simultaneously, without the need to inoculate with pathogens. Pyramiding of resistance genes to provide durable resistance is therefore greatly facilitated. When resistance genes are transferred from wild relatives into a cultivated crop, molecular markers can assist in selecting against the undesired genetic background of the donor parent
| [8] | Ejeta, G. H. (2007). Breeding for Striga Resistance in Sorghum: Exploitation of an Intricate Host–Parasite Biology. Crop science 47(S3), S216–S227. |
[8]
.
According to
| [25] | Nasreldin Mohemed Ahmed (2018): The role of strigolactones in resistance, tolerance and control of Striga infection in Sorghum; PhD Thesis Dissertation submitted in fulfillment of the requirements for the degree of doctor at Wageningen University. |
| [28] | Temesgen Teressa, (2019). “Review on Striga Distribution, Infestation and Genetic Potential in Ethiopian Sorghum (Sorghum Bicolor (L.) Moench)” International Journal of Research Studies in Agricultural Sciences (IJRSAS), 5(2), pp. 23-31. |
[25, 28]
, the application of marker-assisted selection is particularly advantageous when:
1) Resistance tests are difficult, complex, expensive or unreliable;
2) The pathogen is quarantined;
3) Breeding materials are advanced in off-season nurseries where the disease does not occur;
4) Resistance genes are recessive, restricting the effectiveness of back-cross schemes.
Striga resistance breeding in cereals is one case in point. Efforts are currently underway to identify and map genes for qualitative and quantitative resistance to striga in three sorghum mapping populations. These were from three crosses: SRN 39_Shanqui Red; IS 9830_E 36-1; and N13_E 36-1
| [27] | Haussmann, B. I. G., Hess, D. E. Weiz, H. G. and Geiger, H. H. 2000. Improved methodologies for Breeding, Striga resistant sorghums, Field Crops Research. |
[27]
. The identification of individual genes or QTL for striga resistance and their transfer into adapted cultivars will also allow to evaluate whether there are “costs of striga resistance”,
i.e., whether resistance is associated with any yield drag. Such costs of resistance might have been another reason for the slow breeding process in the past.
3.3. Genetic Engineering
Genetic engineering permits the transfer of resistance genes from any organism into a chosen crop. In the case of striga resistance, the main limitation at present is the lack of well-defined resistance genes. However, there is an alternative means by which genetic engineering can be brought to bear on the striga problem. To achieve immediate, cost-effective selective control of parasitic weeds by herbicides,
| [20] | Saunders, A. (1933). Studies in phenorogamic parasitism with particular reference to Striga lutea (Lour). South Africa Department of Agriculture, Science Bulletin, 128, 1-57. |
| [21] | Parker, C. (1984). The influence of Striga species on sorghum under varying nitrogen fertilization. In: Parker, C. Musselman, L. J., Polhill, R. M. and Wilson, A. K. (eds.) Proceedings, 3rd International Symposium on Parasitic Weeds. Aleppo, Syria, 1984. ICARDA, Aleppo. pp. 90-98. |
[20, 21]
proposed the introduction of transgenic, herbicide tolerant crops. According to the above-cited authors, herbicide tolerance in crops affected by parasitic weeds has several positive properties: (1) it allows the control of the parasitic weeds at a very low dosage; (2) it is effective against all major species or strains of the parasite; and (3) it supports or even replaces cultivation methods for control of other weeds. Furthermore, herbicide tolerance should only be used in crops which do not crossbreed with related weeds in the same locality.
The transfer of the XA-17 gene into sorghum could therefore be recommended only for regions, where the crop does not have feral or weedy relatives,
i.e., in Asia, but not in Africa. Even if this condition is respected, there exists the strong possibility of evolution of herbicide resistance in parasitic weeds. The high natural frequency of such mutations and the huge seed output of striga only serve to exacerbate this risk
| [21] | Parker, C. (1984). The influence of Striga species on sorghum under varying nitrogen fertilization. In: Parker, C. Musselman, L. J., Polhill, R. M. and Wilson, A. K. (eds.) Proceedings, 3rd International Symposium on Parasitic Weeds. Aleppo, Syria, 1984. ICARDA, Aleppo. pp. 90-98. |
[21]
. Another consideration involving herbicide-tolerant crops as components of integrated striga control strategies is the ability of farmers to purchase improved seed and the herbicide.
3.4. Breeding for Improved Integrated Striga Control
In addition to selection for host plant resistance, sorghum breeders could consider selecting cultivars for specific adaptation to integrated striga management regimes. For example, the interaction between local sorghum cultivars and fertilizer application or intercropping with legumes could be studied with the aim of selecting cultivars with the highest positive interaction with these measures for grain yield and striga suppression. Another possibility would be to select legume cultivars that effectively induce suicidal germination of
S. hermonthica | [19] | Berner, D. K., Winslow, M. D., Awad, A. E., Cardwell, K. F., Mohn Raj. D. R., & Kim. S. K. (1997). Striga Research Methods - A manual, 2nd edition. International Institute of Tropical Agriculture, PMB 5320, Ibadan, Nigeria. Pp 1-80. |
[19]
. Rotations with legumes increase soil nitrogen and organic matter, and hence enhance the biological control of striga (soil suppressiveness). The mentioned authors identified substantial variation in striga stimulant production among soybean cultivars using a simple laboratory assay. Field trials validated results from laboratory assays, showing reduced parasite emergence and increased cereal yields following rotations with high-stimulant producing legume cultivars
| [8] | Ejeta, G. H. (2007). Breeding for Striga Resistance in Sorghum: Exploitation of an Intricate Host–Parasite Biology. Crop science 47(S3), S216–S227. |
| [9] | Ejeta, G. & Gressel, J. (2007) Integrating New Technologies for Striga Control: Towards Ending the Witch-Hunt. Singapore; Hackensack, NJ: World Scientific. |
| [19] | Berner, D. K., Winslow, M. D., Awad, A. E., Cardwell, K. F., Mohn Raj. D. R., & Kim. S. K. (1997). Striga Research Methods - A manual, 2nd edition. International Institute of Tropical Agriculture, PMB 5320, Ibadan, Nigeria. Pp 1-80. |
[8, 9, 19]
.
Future prospects
Background studies on Striga spp life cycle made some significant advances in the understanding of the biology and physiology of Striga spp. such as after- ripening, germination, haustorial initiation, attachment, penetration and establishment of parasite.However, continued effort is needed in the laboratory and in the field researches to gain a better understanding of factors influencing the different stages of the parasite life cycle so as to develop Striga resistant varieties of sorghum via integrated breeding approach that will lead to effective, economically feasible and environmentally sound.