Growth and body size of mammals are commonly correlated with many life history strategies, including those related to survival and reproduction. However, in certain circumstances suboptimal growth rates and smaller size may be advantageous and adaptive. The water vole Arvicola amphibius is a large vole, about three times the size of a field vole Microtus agrestis, but with similar ecological and reproductive characteristics. Island populations were studied on the coast of northern Norway, just below the Arctic Circle, during 2003-2018, by capture-mark-recapture. The main aims were to study growth rates, asymptotic weight and survival, expecting that the “optimal” weight for surviving the winter would be 140-160 g. The smallest juveniles caught weighed only 21 g and were assumed, based on data from the literature, to be around 14 days old. This age was used as starting point for the growth curve. This, however, may have been about one week too early, as juveniles are more likely not fully weaned and leave their nest of birth until 30-40 g. Initial growth rates in juveniles was relatively high but declined from around 100-120 g or 40-50 days old. The asymptotic weight was not clearly defined, but its maximum was around 150-160 g. Most juveniles that survived the winter weighed between 100 and 160 g in their first summer. Large individual variations in growth rates were found. Overwintered subadults in spring weighed about the same as juveniles did in the autumn but grew quickly in April and May to reach adult size. A specific “optimal” weight for juveniles that survived the winter was not found. The range could be given as 100-160 g, too broad to define an “optimal” weight range. However, those that survived tended to be slightly heavier than those that died. Reproducing adults generally weighed 180-220 g and did not reduce their weight toward the autumn, i.e., to increase winter survival, but very few adults survived even the summer and almost none survived their second winter. Juveniles postponed reproduction until next spring, most likely to take advantage of fresh vegetation growth and less competition.
| Published in | American Journal of BioScience (Volume 12, Issue 3) |
| DOI | 10.11648/j.ajbio.20241203.11 |
| Page(s) | 80-89 |
| Creative Commons |
This is an Open Access article, distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution and reproduction in any medium or format, provided the original work is properly cited. |
| Copyright |
Copyright © The Author(s), 2024. Published by Science Publishing Group |
Growth Rates, “Optimal” Weight, Survival, Voles, Weight Adjustment
Juveniles | Subadults & adults | ||||||
|---|---|---|---|---|---|---|---|
Weight class (g) | Growth rate | a) All caught | b) Re-captured | c) Over-wintered | Growth rate | d) All caught | |
1 | 21-40 | 2.46±0.63 | 5.6 | 3.0 | 0 | ||
2 | 41-60 | 2.83±4.01 | 7.3 | 6.3 | 6.4 | ||
3 | 61-80 | 1.48±1.70 | 13.8 | 12.6 | 5.0 | ||
4 | 81-100 | 1.06±2.38 | 18.7 | 13.2 | 10.6 | 0.1 | |
5 | 101-120 | 0.58±3.83 | 22.4 | 20.4 | 24.1 | 1.78±6.84 | 3.0 |
6 | 121-140 | 0.26±3.07 | 21.6 | 26.1 | 28.4 | 1.45±6.00 | 7.2 |
7 | 141-160 | -0.25±5.47 | 8.8 | 13.2 | 22.0 | 0.37±6.74 | 11.8 |
8 | 161-180 | -2.25±6.66 | 1.4 | 5.1 | 3.5 | 1.14±4.75 | 18.3 |
9 | 181-200 | - | 0.4 | 0 | 0 | -0.31±6.24 | 28.7 |
10 | 202-220 | -2.09±8.36 | 18.1 | ||||
11 | >221 | -2.69±7.36 | 12.7 | ||||
N | 333 | 1870 | 333 | 141 | 277 | 1047 | |
| [1] | Peters, R. H. The ecological implications of body size. Cambridge: Cambridge University Press; 1986. |
| [2] | Case, T. J. Evolution and adaptive significance of postnatal growth rates in terrestrial vertebrates. Q. Rev. Biol. 1978, 53, 243–282. |
| [3] | Cuthill, I., Houston. A. Managing time and energy. In Behavioural ecology: An evolutionary approach, Krebs, J., Davis, N., Eds. Oxford: Blackwell Scientific Publications; 1997, pp. 97–120. |
| [4] | Blanckenhorn, W. U. The evolution of body size: what keeps organisms small? Q. Rev. Biol. 2000, 75, 385–407. |
| [5] | Iverson, S. L, Turner B. N. Winter weight dynamics in Microtus pennsylvanicus. Ecology. 1974, 55, 1030–1041. |
| [6] | Boonstra, R., Krebs, C. J. Viability of large-sized and small-sized adults in fluctuating vole populations. Ecology. 1979, 60, 567–573. |
| [7] | Hansson, L. Ultimate factors in the winter weight depression of small mammals Mammalia. 1990, 54, 397-404. |
| [8] | Hansson, L Size dimorphism in microtine rodent populations: characteristics of growth and selection against large-sized individuals. J. Mammal. 1995, 76, 867–872. |
| [9] | Lidicker, W. Z., Ostfeld, R. S. Extra-large body size in California voles: causes and fitness consequences. Oikos. 1991, 61, 108–121. |
| [10] | Norrdahl, K., Korpimäki, E. Changes in individual quality during a 3-year population cycle of voles. Oecologia. 2002, 130, 239–249. |
| [11] | Mangel, M., Stamps, J. Trade-offs between growth and mortality and the maintenance of individual variation in growth. Evol Ecol Res. 2001, 3, 583–593. |
| [12] | Burthe, S. J., Lambin, X., Telfer, S., Douglas, A, Beldomenico, P., Smith, A., Begon, M. Individual growth rates in natural field, Microtus agrestis, populations exhibiting cycling population dynamics. Oecologia. 2010, 162, 653-661. |
| [13] | Lessels, C. M. 1991. The evolution of life histories. In Behavioural ecology: An evolutionary approach, Krebs, J. R, Davis, N. B., Eds. Oxford: Blackwell Scientific Publications; 1991, pp. 32-68. |
| [14] | Ergon, T., Lambin, X., Stenseth, N. C. Life-history traits of voles in a fluctuating population respond to the immediate environment. Nature. 2001, 411, 1043–1045. |
| [15] | Krebs, C. J. Review of the Chitty hypothesis of population regulation. Can. J. Zool. 1978, 56, 2463–2480. |
| [16] | Oli, M. K. The Chitty effect: A consequence of dynamic energy allocation in a fluctuating environment. Theor. Popul. Biol. 1999, 56, 293–300. |
| [17] | Ergon, T., Speakman, J. R., Scantlebury, M., Cavanagh, R., Lambin, X. Optimal body size and energy expenditure during winter: why are voles smaller in declining populations? Am. Nat. 2004, 165, 442-457. |
| [18] | Wan, X., Zhang, X., Wang, G., Chen, L. Optimal body weight of Brandt’s voles for winter survival. J. Arid. Environ. 2014, 103, 31-35. |
| [19] | Strachan, R. Water vole conservation handbook. Stafford: George Street Press Ltd; 1998. |
| [20] | Frafjord, K. Population dynamics of an island population of water voles Arvicola amphibius (Linneaus, 1758) with one major predator, the eagle owl Bubo bubo (Linneaus, 1758), in northern Norway. Polar Biology. 2022, 45, 1-12. |
| [21] | Zejda, J. The weight growth of the water vole (Arvicola terrestris) under natural conditions. Folia Zool. 1992, 41, 213-219. |
| [22] | Frafjord, K. Influence of reproductive status: home range size in water voles (Arvicola amphibius). PloS ONE. 2016, 11: e0154338 0154331-0154313. |
| [23] | Frafjord, K. Grazing by sheep Ovis aries reduces island populations of water voles. Fauna norv. Serie A. 2014, 34, 79-81. |
| [24] | Frafjord, K. Do eagle owls select larger water voles? Fauna norv. Serie A. 2003, 23, 42-47. |
| [25] | Stoddart, D. M. Breeding and survival in a population of water voles. J. Animal Ecol. 1971, 40, 487-494. |
| [26] | Reichstein, H. Arvicola terrestris (Linneaus, 1758) – Schermaus. In Handbuch der Säugetiere Europas. Band 2/I Rodentia II, Niethammer, J., Krapp, F., Eds., Wiesbaden: Akademische Verlagsgesellshcaft; 1982, pp. 217-252. |
| [27] | Woodroffe, G. L. The water vole. London: The Mammal Society; 1996. |
| [28] | Nazarova, G. G., Evsikov, V. I. Effect of mother's physical condition during pregnancy and lactation on postnatal growth and reproductive success of offspring in water vole Arvicola terrestris. Develop. Physiol. 2008, 39, 100-107. PMID: 18669294. |
| [29] | Nazarova, G. G. Effects of seasonal, ontogenetic, and genetic factors on lifespan of male and female progeny of Arvicola amphibius. Frontiers in Genetics. 2013, 4, Article 100. |
| [30] | Ventura, J., Gosálbez, J. Reproduction potential of Arvicola terrestris (Mammalia, Rodentia) in the Northeast Peninsula Iberian. Zool. Anz. 1990a, 225, 45-54. |
| [31] | Neyland, P. J. Habitat, home range, diet and demography of the water vole (Arvicola amphibious): Patch-use in a complex wetland landscape. Ph.D. thesis, Swansea University, 2011. http://cronfa.swan.ac.uk/Record/cronfa42744 |
| [32] | Capreolus Wildlife Consultancy. The ecology and conservation of water voles in upland habitats. Scottish Natural Heritage Commissioned Report; 2005, No. 099 (ROAME No. F99AC320). |
| [33] | Jeppson, B. Behavioural ecology of the water vole, Arvicola terrestris, and its implications to theories of microtine ecology. Ph. D. thesis, University of Lund, Sweden, 1987. |
| [34] | Ventura, J., Gosálbez, J. Reproductive cycle of Arvicola terrestris (Rodentia, Arvicolidae) in the Aran Valley, Spain. Z. Säugetierk. 1990b, 55, 383-391. |
| [35] | Moorhouse, T. P., Gelling, M., Macdonald, D. W. Effects of forage availability on growth and maturation rates in water voles. J. Animal Ecol. 2008, 77, 1288-1295. |
| [36] | Rogov, V. G., Potapov, M. A., Evsikov, V. I. Sexual structure of the water vole population Arvicola terrestris (Rodentia, Cricetidae) in Western Siberia. Zool. Zhurnal. 1999, 78, 985-XVI. |
| [37] | Semb-Johansson, A., Engh, K. E., Østbye, E. Reproduction, litter size and survival in a laboratory strain of the Norwegian lemming (Lemmus lemmus). In The biology of lemmings, Stenseth, N. C, Ims, R. A., Eds, London: The Linnaean Society of London; 1993, pp. 329-337. |
| [38] | Stenseth, N. C., Ims, R. A. Food selection, individual growth and reproduction—an introduction. In The biology of lemmings, Stenseth, N. C., Ims, R. A., Eds, London: The Linnaean Society of London; 1993, pp 263–280. |
APA Style
Frafjord, K. (2024). No Optimal Weight to Survive the Winter in a Northern Island Population of Water Voles Arvicola amphibius. American Journal of BioScience, 12(3), 80-89. https://doi.org/10.11648/j.ajbio.20241203.11
ACS Style
Frafjord, K. No Optimal Weight to Survive the Winter in a Northern Island Population of Water Voles Arvicola amphibius. Am. J. BioScience 2024, 12(3), 80-89. doi: 10.11648/j.ajbio.20241203.11
AMA Style
Frafjord K. No Optimal Weight to Survive the Winter in a Northern Island Population of Water Voles Arvicola amphibius. Am J BioScience. 2024;12(3):80-89. doi: 10.11648/j.ajbio.20241203.11
@article{10.11648/j.ajbio.20241203.11,
author = {Karl Frafjord},
title = {No Optimal Weight to Survive the Winter in a Northern Island Population of Water Voles Arvicola amphibius
},
journal = {American Journal of BioScience},
volume = {12},
number = {3},
pages = {80-89},
doi = {10.11648/j.ajbio.20241203.11},
url = {https://doi.org/10.11648/j.ajbio.20241203.11},
eprint = {https://article.sciencepublishinggroup.com/pdf/10.11648.j.ajbio.20241203.11},
abstract = {Growth and body size of mammals are commonly correlated with many life history strategies, including those related to survival and reproduction. However, in certain circumstances suboptimal growth rates and smaller size may be advantageous and adaptive. The water vole Arvicola amphibius is a large vole, about three times the size of a field vole Microtus agrestis, but with similar ecological and reproductive characteristics. Island populations were studied on the coast of northern Norway, just below the Arctic Circle, during 2003-2018, by capture-mark-recapture. The main aims were to study growth rates, asymptotic weight and survival, expecting that the “optimal” weight for surviving the winter would be 140-160 g. The smallest juveniles caught weighed only 21 g and were assumed, based on data from the literature, to be around 14 days old. This age was used as starting point for the growth curve. This, however, may have been about one week too early, as juveniles are more likely not fully weaned and leave their nest of birth until 30-40 g. Initial growth rates in juveniles was relatively high but declined from around 100-120 g or 40-50 days old. The asymptotic weight was not clearly defined, but its maximum was around 150-160 g. Most juveniles that survived the winter weighed between 100 and 160 g in their first summer. Large individual variations in growth rates were found. Overwintered subadults in spring weighed about the same as juveniles did in the autumn but grew quickly in April and May to reach adult size. A specific “optimal” weight for juveniles that survived the winter was not found. The range could be given as 100-160 g, too broad to define an “optimal” weight range. However, those that survived tended to be slightly heavier than those that died. Reproducing adults generally weighed 180-220 g and did not reduce their weight toward the autumn, i.e., to increase winter survival, but very few adults survived even the summer and almost none survived their second winter. Juveniles postponed reproduction until next spring, most likely to take advantage of fresh vegetation growth and less competition.
},
year = {2024}
}
TY - JOUR T1 - No Optimal Weight to Survive the Winter in a Northern Island Population of Water Voles Arvicola amphibius AU - Karl Frafjord Y1 - 2024/06/03 PY - 2024 N1 - https://doi.org/10.11648/j.ajbio.20241203.11 DO - 10.11648/j.ajbio.20241203.11 T2 - American Journal of BioScience JF - American Journal of BioScience JO - American Journal of BioScience SP - 80 EP - 89 PB - Science Publishing Group SN - 2330-0167 UR - https://doi.org/10.11648/j.ajbio.20241203.11 AB - Growth and body size of mammals are commonly correlated with many life history strategies, including those related to survival and reproduction. However, in certain circumstances suboptimal growth rates and smaller size may be advantageous and adaptive. The water vole Arvicola amphibius is a large vole, about three times the size of a field vole Microtus agrestis, but with similar ecological and reproductive characteristics. Island populations were studied on the coast of northern Norway, just below the Arctic Circle, during 2003-2018, by capture-mark-recapture. The main aims were to study growth rates, asymptotic weight and survival, expecting that the “optimal” weight for surviving the winter would be 140-160 g. The smallest juveniles caught weighed only 21 g and were assumed, based on data from the literature, to be around 14 days old. This age was used as starting point for the growth curve. This, however, may have been about one week too early, as juveniles are more likely not fully weaned and leave their nest of birth until 30-40 g. Initial growth rates in juveniles was relatively high but declined from around 100-120 g or 40-50 days old. The asymptotic weight was not clearly defined, but its maximum was around 150-160 g. Most juveniles that survived the winter weighed between 100 and 160 g in their first summer. Large individual variations in growth rates were found. Overwintered subadults in spring weighed about the same as juveniles did in the autumn but grew quickly in April and May to reach adult size. A specific “optimal” weight for juveniles that survived the winter was not found. The range could be given as 100-160 g, too broad to define an “optimal” weight range. However, those that survived tended to be slightly heavier than those that died. Reproducing adults generally weighed 180-220 g and did not reduce their weight toward the autumn, i.e., to increase winter survival, but very few adults survived even the summer and almost none survived their second winter. Juveniles postponed reproduction until next spring, most likely to take advantage of fresh vegetation growth and less competition. VL - 12 IS - 3 ER -
The Arctic Museum of Norway, UiT the Arctic University of Norway, Tromsø, Norway
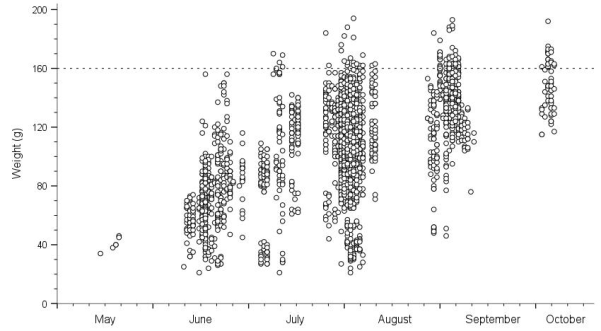
Figure 1. Weight of juvenile water voles across the season, including recaptures. Overall trapping effort differed between months.
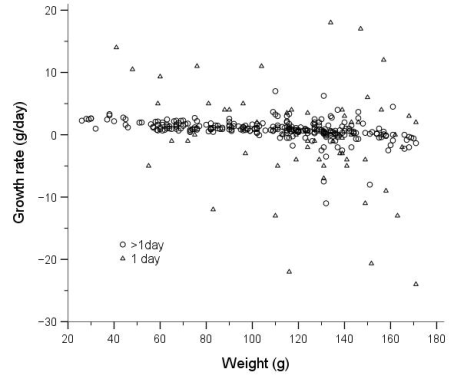
Figure 2. Growth rates of juvenile water voles plotted against body weight, with those recaptured the next day indicated by triangles.
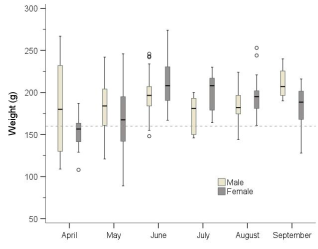
Figure 3. Boxplot of the weight of subadult and adult water voles by sex and month. The dashed line represents 160 g.
Information

