1. Introductory Backgrounds
Growing up in Charleston, South Carolina, the young inquiring Ernest Everett Just (1883–1941) developed an early interest in the biology of marine life. At the age of 13, his mother enrolled him in “Colored Normal Industrial Agricultural and Mechanical College of South Carolina” hoping that he could enter into the teaching profession. Due to concerns over the inferiority of black educational institutions, at the age of 16 he transferred to Kimball Union Academy in Meriden, New Hampshire, and upon graduation, enrolled at Dartmouth University. While a college student, he developed an academic interest in embryology and carried out independent research studies with Professor William Patten (1861-1932). Receiving honors in zoology, with distinctions in botany, history and sociology, he graduated class of 1907, magna cum lauda (the highest distinction that year), with election into Phi Beta Kappa. Thereupon, he began teaching at Howard University in Washington, District of Columbia, first English and rhetoric in 1907, and then biology in 1909, assuming the role of department chair the following year. Through personal introduction, Just was invited by the University of Chicago, Zoology Department chairman Frank Lillie (1870–1947) to join him as an assistant at the Marine Biological Laboratory (MBL) at Woods Hole, Massachusetts. Beginning research in 1909, and while continuing his teaching duties at Howard University, Just was able to earn his PhD remotely from the University of Chicago in 1916, the first African-American to be awarded a degree from that department. His dissertation was entitled “The Mechanics of Fertilization,” and by the time he completed it, he had already published several research articles, either as a co-author with Lillie or as a single author. In 1924, he co-authored a textbook
General Cytology (Univ. Chicago Press) along with Edmund Beecher Wilson (1856-1939), Thomas Hunt Morgan (1866-1945), who later won the Nobel prize, Clarence Erwin McClung (1870-1946) and Margaret Reed Lewis (1881-1970). Sometime thereafter in 1929, Just traveled to Naples, Italy, to carry out experiments at the Anton Dohrn Laboratory, and the following year in 1930, was invited to the Kaiser-Wilhelm-Institut für Biologie in Berlin-Dahlem, Germany, the first American citizen to receive such an invitation, where he participated in laboratory work with a number of Nobel laureates. He continued to travel to Europe with ten more trips up to the year 1938, although after Nazi activities arose in Germany in 1933, he moved his research to Paris, France, and the Roscoff Station Biologique along the English channel. When Germany invaded France in 1940, he was drafting a paper “Unsolved Problems in General Biology” and was briefly interred in a prison camp; however, through the help of his partner (and later co-author) Hedwig Schnetzler (1907-1907), a philosophy student whom he married in Berlin, he was soon released and returned to the United States in September 1940. When he arrived home, he was shortly thereafter diagnosed with pancreatic cancer and died in October 1941. During his prolific and short, but brilliant, career, he published two textbooks and over seventy original research articles, which can be found in such journals as:
American Journal of Physiology,
American Naturalist,
Anatomical Record,
Biological Bulletin,
Cytologia,
Natur-wissenschaften,
Physiological Zoology,
Protoplasma,
Science, Wilhelm Roux’s
Archiv für Entwicklungsmechanik,
Der Organismen (now
Development, Genes and Evolution), and
Zeitschrift für Zellforschung und Mikroscopik Anatomie | [6] | Byrnes, W. M., Eckberg, W. R. (2006) Ernest Everett Just (1883–1941)—An Early Ecological Developmental Biologist. Developmental Biology, 296(1), 1-11. https://doi.org/10.1016/j.ydbio.2006.04.445 |
| [55] | Manning, K. R. (1985) Black Apollo of Science: The Life of Ernest Everett Just. Oxford press. |
[6, 55]
.
Born a generation later but during roughly the same time period, William Thomas Fontaine (1909-1968) had precocious aspirations to mathematics and philosophy as a youth. Growing up in Chester, Pennsylvania, he was able to enter the integrated Chester High School, where the academic course of study required four years of English, three of mathematics, three of history, three of foreign language and two of science. However, upperclassmen were allowed the option of carrying a ‘fifth major,’ which Fontaine took in mathematics and Latin. Outside of academics, he also played for the Chester High football team and participated in the extra-curricular Dunbar Literary Society, named in honor of African-American poet Paul Laurence Dunbar (1872–1906), whose works have been noted for their “philosophical musings” on literary characters who “embodied the demise of individualism in the face of hereditary and environmental determinism, and therefore […] lack control over the strong forces, internal or external, that cause a visceral human effect” (
| [30] | Jarrett, G. A. (2022) Paul Laurence Dunbar: the Life and Times of a Caged Bird. Princeton Press. |
[30]
, p. 321). After graduation, Fontaine entered college at Lincoln University, located about 30 miles west of his home city. He began writing for the school newspaper, with analytical essays, philosophical reflections and poems; in these, he contended that African-Americans needed to show “ability, aggressiveness, and cooperation” (
| [52] | Kuklick, B. (2011) Black Philosopher, White Academy: The Career of William Fontaine. University of Pennsylvania Press, Philadelphia. |
[52]
, p. 18) to succeed. Fontaine graduated first in his class in 1930, and for the following six years, taught Latin, history and government at Lincoln University, while he pursued an MA in philosophy from the University of Pennsylvania; he completed this degree in 1932 and then went on to earn his PhD in philosophy from the same university in 1936. His dissertation was entitled “The Concept of Fortune in Boethius and Giordana Bruno”
| [19] | Fontaine, W. T. (1936) Fortune, Matter and Providence: A Study of Ancius Severinus Boethius and Giordano Bruno. PhD dissertation, University of Pennsylvania. https://philpapers.org/rec/FONFMA |
[19]
. Upon graduating, Fontaine married an acquaintance from Philadelphia in 1936, Willabelle Hatton (?-?) from Iva, South Carolina. From 1936 to 1942, he taught as professor of philosophy and history at Southern University in Baton Rogue, Louisiana. In 1946, Fontaine enlisted in the U.S. Army, and also became philosophy department chair at Morgan State College in Baltimore, Maryland, the same year. In 1947, he was visiting lecturer at University of Pennsylvania; then in 1949, he was appointed assistant professor; and finally in 1963, he was made associate professor, the first full tenured African-American faculty member at the university. In 1968, Fontaine died from health complications developed from a brief tuberculosis infection contracted in 1949. He is remembered for his book
Reflections on Segregation, Desegregation, Power and Morals (1967), as well as scholarly articles on diverse topics in
The Journal of Philosophy,
American Journal of Sociology,
Philosophy and Phenomenological Research,
Journal of Negro History,
Présence Africaine,
Philosophy of Science, and
Southern University Bulletin | [52] | Kuklick, B. (2011) Black Philosopher, White Academy: The Career of William Fontaine. University of Pennsylvania Press, Philadelphia. |
[52]
.
2. Ernest Everett Just, Jacques Loeb, & the Corrective Factors to 1-D Biology
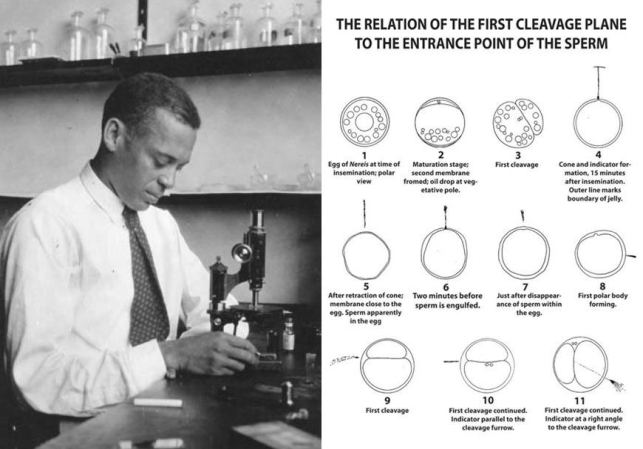
The early developmental biology papers of E.E. Just were of an essential mathematical nature. Carrying out studies of embryonic development, he recorded the correlation between the point of sperm entry and the plane of blastomere cell development, with two major results: (1) symmetries of blastomere division are variable from species to species, i.e. symmetry-breaking in evolutionary history; and (2) symmetries of blastomere division begin to vary in the course of development, i.e. symmetry-breaking during embryonic growth. For example, his first paper, “The Relation of the First Cleavage Plane to the Entrance Point of the Sperm” published in 1912 in
Biological Bulletin,
recorded the relationship in marine Nereis [marine worms] between the embryonic cleavage plane [where a fertilized egg divides into blastomere cells] and the point of sperm entrance, and found between 50% and 95% coincided (at left-right, 180°) (
| [34] | Just, E. E. (1912) The Relation of the First Cleavage Plane to the Entrance Point of the Sperm. The Biological Bulletin, 22 (4), 239-252. https://doi.org/10.2307/1535889 |
[34]
, p. 245). However, as he noted “there are other [species’] eggs in which the future median plane does not fall in the plane of the first cleavage,” including Crepidula (snails), where “the first cleavage plane is at right angles to the future median plane” (at up-down, 90°), and Claetopterus (feeder worms) where “the axis of the first cleavage spindle lies in the longitudinal axis of the embryo” (at forward backwards), and Amia (bowfish) “in which coincidence with any cleavage plane is wanting” (any direction) (
| [34] | Just, E. E. (1912) The Relation of the First Cleavage Plane to the Entrance Point of the Sperm. The Biological Bulletin, 22 (4), 239-252. https://doi.org/10.2307/1535889 |
[34]
, p. 250). In 1915, Just extended this study to Platynereis megalops (marine worm) and found that “the figures show that there is no constant disposition of granules at the point of sperm entry-certainly nothing of the nature of a cone, as in Nereis” (
| [35] | Just, E. E. (1915) An Experimental Analysis of Fertilization in Platynereis Megalops. The Biological Bulletin, 28 (2), 93-114. https://doi.org/10.2307/1536272 |
[35]
, p. 228). In the following decade, he published a two-part study “Studies Of Cell Division: I” (1922)
| [38] | Just, E. E. (1922a) Studies of Cell Division: I. The Effect of Dilute Sea-water on the Fertilized Egg of Echinarachnius Parma during the Cleavage Cycle. American Journal of Physiology, 61 (3), 505-515. https://doi.org/10.1152/ajplegacy.1922.61.3.505 |
[38]
and “Studies Of Cell Division: II” (1928)
| [40] | Just, E. E. (1928) Studies of Cell Division. II. The Period of Maximum Susceptibility to Dilute Sea-water in the Cleavage Cycle of the Fertilized Egg of Arbacia. Physiological Zoology, 1(1), 26-36. https://doi.org/10.1086/physzool.1.1.30151032 |
[40]
comparing the cleavage-plane formation of the Echinarachnius Parma (sand dollar) with the Arbacia (sea urchin), finding that both regulated cell-division patterns by the formation of hyaline-plasm layer that modulated fluid susceptibility into the elongating ova cells. Later, he summarized some of the general ideas of these studies in a 1937 paper; for example:
“Eggs of most bilaterally symmetrical animals begin their development as radially symmetrical [i.e., cylindrical symmetry] structures and there fore show a polar axis. But at the moment after fertilization when bilaterality [i.e., left-right symmetry] appears in such an egg, we can no longer speak of an axis. In a bilaterally symmetrical organism—egg or adult —there exists no line common to planes as in a radially symmetrical one [i.e., cylindrical symmetry]. Here, accurately speaking, we can use only the term, plane of symmetry” (
| [44] | Just, E. E. (1937) Phenomena of Embryogenesis and Their Significance for a Theory of Development and Heredity. The American Naturalist, 71(733), 97-112. https://doi.org/10.1086/280711 |
[44]
, p. 102).
Figure 1. (L) Ernest Everett Just in 1925 photo, University of Chicago archives; (R) Just’s drawings of the cleavage plane during development in worm Nereis, from Biol Bull (1915): 240-249.
The distinct biological processes of symmetry breaking found between different species in evolutionary history and within a species’ embryonic development required a mathematical approach of descriptive geometry. Moreover, the analysis of the biochemical and biophysical factors that underlied these distinctive geometric patterns, according to Just’s research, were mutli-factorial. Only by first conducting experiments on a variety of species and introducing modifications at a variety of times could the systematic organizational processes be clarified. Then, with these insights, a quantized biology could be detailed for a variety of developmental patterns between and within species.
Reflections on these complex, interlocked patterns most certainly led Just to perhaps his most original contribution to the general theory of biology, i.e., cytoplasmic potencies being restricted by genetic expression due to a series of reciprocal interactions between the cytoplasm and nucleus. However, his theory went against the mainstream consensus view of the nucleus as a control center for cellular activity. In fact, many at the time had begun to consider these developmental processes hard-programmed by the cell nucleus. Initial statements of this mechanistic nuclear theory were at first only implicitly murmured. For example, Edwin G. Conklin (1908) stated: “[T]he mechanism of heredity is merely the mechanism of differentiation. The peculiarity which distinguishes the differentiation of the egg cell from that of any tissue cell ‒ is the fact that the latter gives rise only to a particular type of cell, with the formation of which differentiation comes to an end, whereas the former undergoes a long series of differentiations and gives rise to a complicated organism” (
, pp. 90-91). Similarly, Thomas Hunt Morgan (1917) wrote: “[E]ven if we postulated nothing more about them than [genetic factors’] independence of each other and their distribution in the germ cells, we could still handle the Mendelian results on a purely mathematical basis that would enable us to predict what new combinations should give” (
, p. 515). These initially passively stated ideas, however, became increasingly a dominant view held by Just’s fellow biologists
| [26] | Gilbert, S. F. (1988) Cellular Politics: Ernest Everett Just, Richard B Goldschmidt, and the Attempt to Reconcile Embryology and Genetics. In: American Development of Biology, Eds. R. Ronald et al., De Gruyter, Berlin, pp. 311-346. https://doi.org/10.9783/9781512805789-013 |
[26]
. Again, the statements of Thomas Hunt Morgan (1926) exemplify this trend:
“A change in a gene produces definite effects on the developmental processes. It affects one or more of the characters that appear at some later stage in the individual. In this sense, the theory of the gene is justified without attempting to explain the nature of the causal processes that connect the gene and the characters. Some needless criticism of the theory has arisen from failure to clearly understand this relation. […] [T]he theory has been unfairly criticised on the grounds that the organism is a physico-chemical mechanism while the genetic theory fails to account for the mechanism that is involved. But the only assumptions made [...] involve no assumptions inconsistent with physical principles” (
| [60] | Morgan, T. H. (1926) The Theory of the Gene. Yale Press, New Haven. |
[60]
, pp. 26-27).
As years went by, biologists began to articulate more views along these lines stated by Morgan. For example, H. H. Dixon (1936) wrote that “genes, in the same nucleus, make their presence felt in moulding the structure and metabolism of the cell” (
, p. 615). His statement had a certain emphatic quality that biased his perspective on his research, and this academic pattern of nucleus-centered theories continued again with others. For example, Donald F. Jones (1938):
“The changes that take place during development resulting from the loss or shift of chromosomes or chromosome segments have a direct bearing on the control of growth [...] The evidence derived from genetic and cytological observations shows that changes originate in aberrant chromosome behavior. The exact nature of this aberration is not known in every case, but the final result of these nuclear changes is expressed in the cytoplasm. Since in the single cell mosaics no cell division has taken place following the first alteration in nuclear constitution, the changes in cell size, shape and composition are apparently due to some influence originating in the nucleus and passing through the nuclear membrane” (
, p. 401).
More examples could be collated with these, all of which showing alignment with the view that the nucleus had the primary function of determining patterns of cellular development. To this day, statements can be found in biological literature that attribute titles of prestige to the nucleus such as the “control center”
or the “command center”
| [64] | Pennisi, E. (1998) The Nucleus’s Revolving Door. Science, 279(5354), 1129-1131. https://doi.org/10.1126/science.279.5354.1129 |
| [73] | Vinson, V., Purnell, B. A., Chin, G. J., Marx, J. (2000) Macromolecular Ballet. Science, 288(5470), 1369-1369. https://doi.org/10.1126/science.288.5470.1369 |
[64, 73]
or also the “cellular headquarters”
| [70] | Thakur, P., Nayyar, H. (2013) Facing the Cold Stress by Plants in the Changing Environment: Sensing, Signaling, and Defending Mechanisms. In: Plant Acclimation to Environmental Stress, Eds. N. Tuteja, S. S. Gill, Springer Press, New York, pp. 29-69. https://doi.org/10.1007/978-1-4614-5001-6_2 |
[70]
.
However, Just perceived that these over-emphasized one-dimensional conceptions of the cellular hierarchy overlooked the complex and interdependent functions of embryonic development. Based on his experimental data and that of others, it was evident to him that the cytoplasm was not mere amorphous junk matter, but a detailed structure of cellular potencies that actively interacted with and even controlled nuclear activity. As early as 1929, Just had stated that “The reactivity of the cell as a whole—its individual and peculiar response to stimulation with attendant measurable physical and chemical changes— is largely, if indeed not wholly, a cortical (ectoplasmic) phenomenon” (
| [41] | Just, E. E. (1929) The Production of Filaments by Echinoderm Ova as a Response to Insemination, with Special Reference to the Phenomenon as Exhibited by Ova of the Genus Asterias. The Biological Bulletin, 57(5), 311-325. https://doi.org/10.2307/1537040 |
[41]
, p. 322). However, emphatic statements with terminology such as “largely” and “wholly” were only rarely to be found Just’s earlier writings. Increasingly though, the dominant view of the nucleus as the command center of the cell was permeating through the larger scientific community, and embryologists and developmental biologists perhaps were waiting for further clarification on the growing preponderance of a scholarly trend. Although trained as an experimentalist with numerous original publications in his sub-discipline, Just found another opportunity to address what appeared to him to be an aberrant theoretical emphasis. As he wrote, there was an alternative to the mainstream view (1936):
“This ultra-mechanistic rigidity of the gene-theory renders it valueless for explaining the process of differentiation, a process marked by the egg’s inherent plasticitv and by its mobile responses to external influences [...] They confess ignorance as to how the genes act both in development and in heredity (Morgan, 1924, and elsewhere). They speculate concerning the possibility of gene-control […], never allowing for the far more reasonable possibility that, as unchanging particles that grow, the genes may add to themselves substances from the cytoplasm. Speculation unsupported by fact is often interlarded with experimental data and supposition is supplemented by supposition as the situation demands.” (
| [43] | Just, E. E. (1936) A Single Theory for the Physiology of Development and Genetics. The American Naturalist, 70(728), 267-312. https://doi.org/10.1086/280666 |
[43]
, pp. 290-291).
His assessment of the mainstream gene-centric position was straightforward: that the theory was being propted up by indefensible arguments and that data were being manipulated to fit the theory. While Byrnes and Eckberg (2006) have contended that Just was “incorrect in denying a central role for genes in development” (
, p. 269), other articles have suggested that his alternative theory was prescient of future advances in cell biology. Crow, for example, has noted that contributions as having been “imaginative, but often not fully supported theories,” although perhaps he was really a “pioneer in the new field, eco–devo, in which the emphasis is on the organism as a whole” (
, p. 1740). Moreover, there is now additional evidence that Just’s theory of cytoplasmic factors effecting nuclear reactions has recently received surprising experimental validation by Pagliara et al.
, whose work has shown that chromosomes are actually “sponge-like” in a selective manner, and are able to respond to various cytoplasmic factors that determine how the individual cell later changes into differentiated cell types.
Reviewing these years of his professional activity, Byrnes (2015) has further noted that “after 1936, Just’s papers became increasingly philosophical. This reflected both his desire to apply his ideas about the importance of the cell surface more broadly and his increased willingness to challenge those [who] were too reductionistic” (
, p. 26). His 1937 paper, for example, wrote of “the need for a synthesis of a vast number of particularistic studies and for a [new] biological philosophy” (
| [44] | Just, E. E. (1937) Phenomena of Embryogenesis and Their Significance for a Theory of Development and Heredity. The American Naturalist, 71(733), 97-112. https://doi.org/10.1086/280711 |
[44]
, p. 540), contending that the gene-centric theory had “gone too far in a mechanistic (but in no sense physical) direction and had given rise to unfounded hopes that the grand problem of life had been solved” (
| [44] | Just, E. E. (1937) Phenomena of Embryogenesis and Their Significance for a Theory of Development and Heredity. The American Naturalist, 71(733), 97-112. https://doi.org/10.1086/280711 |
[44]
, p. 540). He further wrote of experiments “so varied that at first thought they seem to defy reduction to a common basis where explanation might be possible” (
| [44] | Just, E. E. (1937) Phenomena of Embryogenesis and Their Significance for a Theory of Development and Heredity. The American Naturalist, 71(733), 97-112. https://doi.org/10.1086/280711 |
[44]
, p. 542) and of his cytoplasmic-centered “hypothesis-[of which] more cannot be proffered in the present state of knowledge-[as being] wholly consistent with the established facts as far as one can reduce them to order” (
| [44] | Just, E. E. (1937) Phenomena of Embryogenesis and Their Significance for a Theory of Development and Heredity. The American Naturalist, 71(733), 97-112. https://doi.org/10.1086/280711 |
[44]
, p. 547). Similar philosophical language permeated his
Biology of the Cell Surface (1939), where he of the “gene-theory of heredity [as] an ultra-mechanical rigorously bound concept” making it “inadmissible as an explanation” and offering “no help in our attempt to explain differentiation” (
| [45] | Just, E. E. (1939a) The Biology of the Cell Surface. P. Blakiston, Philadelphia. |
[45]
, p. 325). He continued by arguing that “the gene-theory as formulated may not be the only way of interpreting the vast amount of reliable data accumulated by the numerous geneticists the world over” (
| [45] | Just, E. E. (1939a) The Biology of the Cell Surface. P. Blakiston, Philadelphia. |
[45]
, p. 325). No longer simply presenting basic data collected from experiments, either as descriptive observations or as quantitative analysis, Just was now venturing to unite his more precise scientific research studies into a radically different theoretical and philosophical side of the scholarly community of biologists. In doing so, it is possible to perceive that he over-emphasized his position, and that he perhaps lost some of his previous tone of experimental and professional rigor. For example, Sapp has summarized: “Just inverted the main propositions of his day about gene and cytoplasm. While it had been proposed that chromatin was at the basis of life’s origin, he reasoned that most likely the ectoplasm came first and that chromatin was made from cytoplasm not the inverse” (
| [67] | Sapp, J. (2009) ‘Just’ in Time: Gene Theory and the Biology of the Cell Surface. Molecular Reproduction and Development, 76(10), 903-911. https://doi.org/10.1002/mrd.21044 |
[67]
, p. 907).
However, his direction of thought at this time was not entirely in the abstract realm of philosophy, and with his earlier background he continued to follow his subfield of scientific investigations. As early as 1919, in fact, Just had begun closely following the experiments of Jacques Loeb (1859-1924) on the formation of hyaline plasma layers in Echinarachnius eggs and noted that, in his own lab, “experiments [were] being made at present with the hope of analyzing farther these cortical responses” (
| [36] | Just, E. E. (1919) The Fertilization Reaction in Echinarachnius Parma. I. Cortical Response of the Egg to Insemination. The Biological Bulletin, 36 (1), 1-10. https://doi.org/10.2307/1536454 |
[36]
, pp. 8-9). Loeb had called his successful initiation of urchin embryonic development “the superficial cytolysis-corrective factor”
. Based on Loeb’s work, and having carried out several experiments, Just wrote in 1920 that Loeb had “definitely commit[ted] himself to the cytolysis theory” caused by living spermatozoon, or by physico-chemical agencies of superficial causes, but that “against this theory of activation several potent objections may be raised” (
| [37] | Just, E. E. (1920) The Fertilization-Reaction in Echinarachnius Parma. IV. A Further Analysis of the Nature of Butyric Acid Activation. The Biological Bulletin, 39 (5), 280-305. https://doi.org/10.2307/1536722 |
[37]
, p. 298). He further had contended that Loeb wrote of cleavage “when he meant disintegration” (
| [37] | Just, E. E. (1920) The Fertilization-Reaction in Echinarachnius Parma. IV. A Further Analysis of the Nature of Butyric Acid Activation. The Biological Bulletin, 39 (5), 280-305. https://doi.org/10.2307/1536722 |
[37]
, p. 299) and that he “conceived ‘artificial parthenogenesis’ as a process comparable to hemolysis and all hemolytic agents as artificial excitants of development” (
| [37] | Just, E. E. (1920) The Fertilization-Reaction in Echinarachnius Parma. IV. A Further Analysis of the Nature of Butyric Acid Activation. The Biological Bulletin, 39 (5), 280-305. https://doi.org/10.2307/1536722 |
[37]
, p 300). Following upon this paper, Just noted that there were “two outstanding features of Loeb’s work” that demonstrated its inability to induce embryonic development, specifically, “by exposing urchins’ eggs to butyric acid in sea-water before or after exposure to hypertonic sea water” (
| [39] | Just, E. E. (1922b) Initiation of Development in the Egg of Arbacia. I. Effect of Hypertonic Sea-water in Producing Membrane Separation, Cleavage, and Top-swimming Plutei. The Biological Bulletin, 43 (6), 384-400. https://doi.org/10.2307/1536595 |
[39]
, p. 384) made no difference. Explaining these failed results, Just contended that “Loeb did not make exact observations with various concentrations of salt” (
| [39] | Just, E. E. (1922b) Initiation of Development in the Egg of Arbacia. I. Effect of Hypertonic Sea-water in Producing Membrane Separation, Cleavage, and Top-swimming Plutei. The Biological Bulletin, 43 (6), 384-400. https://doi.org/10.2307/1536595 |
[39]
, p. 397). Following Loeb’s death in 1924, Just continued research on this topic and described experiments in 1939b that clarified this experimental discrepancy which had permeated the literature for decades: “The method as modified by Heilbrunn and used subsequently by Lillie, Moore and myself for the egg of Arbacia is applicable” (
| [46] | Just, E. E. (1939b) Basic Methods for Experiments on Eggs of Marine Animals. P. Blakiston, Philadelphia. |
[46]
, pp. 46-47). Not forgetting his earlier work, Just sought to add rigorous precision to this study. It has been noted that Loeb had “poor experimental technique” and made “failed attempts—some notoriously involving sloppy experimental techniques” (
| [7] | Byrnes, W. M., Newman S. T. (2014) Errnest Everett Just: Egg and Embryo as Excitable Systems. Journal of Experimental Zoology Part B: Molecular and Developmental Evolution, 322(4), 191-201. https://doi.org/10.1002/jez.b.22567 |
[7]
, p. 10). Continuing his earlier work from the 1920s into this 1939b book, Just focused on these experiments, even as philosophical questions became increasingly voiced in the scientific community.
It is perhaps ironic at this point that as Just had continued to focus his efforts on experimental exactitude, he walked directly into another ‘celebrity culture’ of biological philosophy. Loeb’s earlier experiments which he had taken aim at were considered by some the first experimental evidence of abiogenesis, i.e., the creation of life from non-living matter. Because of the fascinating and consequential nature of such experiments, Loeb was profiled in
McClure’s Magazine in 1902 as a new ‘Doctor Faustus,’ and he lived up to the title when he was recorded with the statement: “I wanted to go to the bottom of things. I wanted to take life in my hands and play with it […] I wanted to handle it in my laboratory as I would any other chemical reaction—to start it, stop it, study it under every condition, to direct it at my will!” (quoted in
| [63] | Pauly, P. (1987) Controlling Life: Jacques Loeb and the Engineering Ideal in Biology. Oxford Press. |
[63]
, p. 102). When Just challenged Loeb’s theories, he inadvertently became the next profiled scientist who had created life from nothing. However, as some science historians have noted: “The study of parthenogenesis is [actually] very old, with major papers appearing on the subject as early as 1774 [...] Several extensive reviews were written during the next century, particularly between 1840 and 1870” (
, p. 43). Moreover, Just was emphatic to note that not all artificial treatments at parthenogenesis led to fully mature offspring:
“Eggs [of platynereis] were therefore treated with KCl, KOH, and NaOH in sea water for various lengths of time and then subjected to heat, shaking, and centrifugal force. In no case did I procure cleavage although the first agent in each case caused maturation. With nereis, on the other hand, KCl and subsequent warming in sea water induces development (see Just 15). It is interesting to note that eggs subjected to heat in the minute quantities of sea-water that permit fertilization do not develop beyond maturation. Apparently,
the conditions for successful artificial initiation of development are more exacting than those for successful insemination” (
| [35] | Just, E. E. (1915) An Experimental Analysis of Fertilization in Platynereis Megalops. The Biological Bulletin, 28 (2), 93-114. https://doi.org/10.2307/1536272 |
[35]
, p. 109).
Clearly, if Just aspired to Loeb’s exaggerated claims of having created life from nothing, he also recognized that greater precision was needed. Additionally, he wrote statements cautioning other scientists against the supposition that any such creations were inevitably possible, e.g., that “we do not know that there is no discontinuity between the non-living and the living world and we certainly possess no evidence for the postulate that living phenomena can be expressed in terms of groupings and displacements of ultimate particles” (
| [45] | Just, E. E. (1939a) The Biology of the Cell Surface. P. Blakiston, Philadelphia. |
[45]
, p. 12). Such then were his basic criticisms of Loeb’s corrective factor theory.
Perhaps what is lost— in this fascinating but drawn-out fiasco with Loeb’s parthenogenetic experiments—was that both of these biologists had been acquiring the habit of using the language of mathematical physics to describe their theoretical process of embryonic differentiation. Loeb’s “superficial cytolysis corrective factor” was certainly a noticeable reference to the Lorentz corrective-factors of electromagnetism and relativity. Examples of this terminology abound in publications from this time period. E.g., “within a small element of volume the space time coordinates are changed by a transformation of the same form as the Lorentz-Einstein, save for the magnification factor λ
-1” (
| [14] | Cunningham, E. (1910) The Principle of Relativity in Electrodynamics and an Extension Thereof. Proceedings of the London Mathematical Society, 2(1), 77-98. https://doi.org/10.1112/plms/s2-8.1.77 |
[14]
, p. 91). E.g. “If the velocity of drift of the electrons of the current is about one centimeter per second, and we deal with corrective terms of the magnitude of those of relativity, the departure from Coulomb's law is given by a factor of about (1+10
-21)” (
, p. 134). E.g., “For the deviation of light in passing the sun it is enough to observe that the equation of motion of the light pulse is to be obtained from the exact equation [...], and its deviation is entirely due to the Einstein correction term.” (
, p. 161). Generally understood, these corrective factors were needed because laws of physics are only valid within a certain domain of applicability; a more general theory might involve more complicated mathematics or perhaps even a different geometrical construction. It is certainly not clear that Just was in any way opposed to the concept of domains of validity in physico-chemical laws and ‘corrective factors.’ He had insightfully noted that the space-time of living matter might be a different domain than the space-time of non-living matter: “Living organization is dynamic whereas the application of chemical analysis by necessity demands destruction of the very space-time structure which is the changing organization characteristic of life” (
| [45] | Just, E. E. (1939a) The Biology of the Cell Surface. P. Blakiston, Philadelphia. |
[45]
, p. 42). However, he also late in his career began to issue statements that suggested he had possibily lost a belief in some of the underlying assumptions of the approaches of mathematical biology: “by some great discovery made in total ignorance of the morphological substratum of biology, someone might be able to appreciate the secret of life in its entirety […] But even if there should be revealed to us the
ultimate space-pattern, there would still remain the problem of the changes of this pattern in time” (
| [45] | Just, E. E. (1939a) The Biology of the Cell Surface. P. Blakiston, Philadelphia. |
[45]
, pp. 25-26). Given such statements, combined with his experimentally-based opposition to Loeb’s “superficial cytolysis-corrective factor,” it is arguable that Just had overlooked new possibilities for groundbreaking advances in mathematical biology. Perhaps he had merely waxed poetic about the ‘secret of life’ to assure the public that artificial life was nowhere on the horizon of biological laboratory work. Or perhaps he had left open questions to a future generation for the possibility of the discovery of the ‘ultimate space-pattern’ in living organisms. But the questions, both about Just’s personal opinions on these topics as well as the mathematical limits inherent to a general theory of biology, still remain.
3. William Fontaine, Samuel Alexander, & the Expanding-Universe of Biology
William T. Fontaine’s 1936 dissertation “The Concept of Fortune in Boethius and Giordana Bruno” certainly has some implicit bearing on the topic of biology and mathematics, although properly it deserves further consideration as such. Boethius’ metaphysics of biological life was profoundly informed by the ancient school of Aristotelian philosophy, and therefore, Fontaine’s association of Boethius (c.480–524) with the Renaissance philosopher Giordano Bruno (1548-1600) might be taken as looking backwards in philosophical history rather than forwards toward philosophical problems on the horizon. This is not to say that the mathematical concepts that Fontaine referenced in his dissertation are obsolete; rather, they belong to a certain time period and school of philosophy. For example:
“Prior to all material things that appear in the life cycle such as ‘stone, earth, cadaver, man, embryo, blood,’ logic demands that there be a material principle from the infinite fecundity of which all things arise. Nor does this material principle exclude or stand irreducible before an opposing formal principle. Both are only different aspects of the same infinite universe for not only may we see all particular forms as arising either from a single ‘perfect species’ or coming forth from the bosom of an infinite matter” (
| [19] | Fontaine, W. T. (1936) Fortune, Matter and Providence: A Study of Ancius Severinus Boethius and Giordano Bruno. PhD dissertation, University of Pennsylvania. https://philpapers.org/rec/FONFMA |
[19]
, p. 5).
Clearly many of the core ideas here predate the development of modern quantitative mathematical biology. Nonetheless other statements are more suggestive; for example: “Bruno’s cosmology and teleology follow from his ontology” meaning, according to Fontaine, that every form of life has innate and perhaps infinite capacities to become every other possible thing, and thus the “mode, regardless of its private aspiration, must live in its constant flux, and every transformation of it contributes toward an actualization” (
| [19] | Fontaine, W. T. (1936) Fortune, Matter and Providence: A Study of Ancius Severinus Boethius and Giordano Bruno. PhD dissertation, University of Pennsylvania. https://philpapers.org/rec/FONFMA |
[19]
, p. 34). Adding more to these ideas, the concept of survival, which can be quantified in evolutionary biology, finds a context in these ancient, medieval and Renaissance philosophies, which Fontaine made keen statements to in his presentation:
“Self-preservation is then a cosmological principle based upon an insufficient concept of the ultimate nature of modal being and of all being. The mode sees only the form in this present setting; but the play goes on behind its back for the life of all modes must consist of a constant influx and efflux of elementary principles ending not in preservation of the particular mode but in loss and transition” (
| [19] | Fontaine, W. T. (1936) Fortune, Matter and Providence: A Study of Ancius Severinus Boethius and Giordano Bruno. PhD dissertation, University of Pennsylvania. https://philpapers.org/rec/FONFMA |
[19]
, pp. 35-36).
The description of self-preservation and survival takes on both a metaphysical aspect and a quantifiable biological aspect – perhaps a unique original contribution by Fontaine. Moreover, many of these terms such as flux, mode, preservation, transition have clear mathematical implications in field equations – also capable of building bridges to ancient metaphysical concepts, as shown in this fascinating text. Profound as his 1936 dissertation was, however, after this publication, much of Fontaine’s research shifted toward sociological and historical questions.
In his 1939 paper about Just’s biological work, Fontaine attributed at least three major features of relevance to Kantian philosophy to be found in his research, and particularly, to be found in the text
Biology of the Cell Surface: (1) the methodology was similar to that of
Critique of Pure Reason (1781/1787); (2) through his embryological experiments, Just had been closer than anyone in artificially creating life, a topic relevant to a number of sections from
Critique of the Power of Judgment (1790); and (3) the mathematical features of Just’s theories had implications beyond 3-dimensional Euclidean geometry and 4-dimensional space-time, ideas which are nowhere to be found in the Kantian corpus. German writers in the 19
th century had taken Kant’s philosophy into an interactive response to Darwin’s theory of evolution by natural selection, exemplified by a number of books, including
Natürliche Schöpfungs-geschichte (1868) by Ernst Haeckel (1834-1919),
Kant und Darwin (1875) by Fritz Schultze (1846-1908),
Studien zur Descendenztheorie (1875) by August Weismann (1834-1914), and others. However, Just had set his work as an alternative to Darwin: “the theory of mutual aid and co-operation may be a better explanation of the cause of evolution than the prevailing popular conception of Darwin’s idea of the struggle for existence […] [T]he means of co-operation and adjustment is the ectoplasm [...] can go farther” (
| [45] | Just, E. E. (1939a) The Biology of the Cell Surface. P. Blakiston, Philadelphia. |
[45]
, p. 367). Therefore, Fontaine’s reading of Just might be considered as a distinct or, perhaps, an even more advanced application of Kantian philosophy to biological theories.
(1) Regarding Just’s method, the general scientific method outlined in the
Critique of Pure Reason entails the idea that analysis and synthesis are two poles of the dynamic process of refining and expanding our knowledge. For example, it is not enough to only analyze a phenomenon into its most simple elements without determining its relation to another phenomenon; one must further discern if both are constituents of a more basic phenomenon or perhaps are related by a law. Similarly, it is not enough to only analyze a law into its most simple statement without determining its relation to another law; one must further discern if both are manifestations of a more basic law or are perhaps are related by an idea. Those who have studied the
Critique in detail could indeed cite passages from A14/B28, A567/B595 and A645/B673
| [47] | Kant, I. (1781/1787 [1998]) Critique of Pure Reason. Trans. & Eds. P. Guyer, A. Wood. Cambridge Press. |
[47]
, for example, and emphasize this reading throughout the entirety of the text. — Review articles provide succinct statements of the essence of it: “the elements of a science are discovered in the analysis somehow, and combined in the synthesis, [to establish] the systematic unity of the elements” (
| [57] | Merritt, M. B. (2004) Drawing from the Sources of Reason: Reflective Self-knowledge in Kant’s First Critique. PhD dissertation, University of Pittsburgh. https://philpapers.org/rec/MERDFT |
[57]
, p. 28); or: “knowledge is a combination of the analytic and synthetic” (
| [58] | Minimah, F. I. (2011) Analytic-Synthetic Dichotomy in Kant and the Logical Positivists: A Comparative Analysis. Journal fo the Department of Philosophy - University of Nigeria, Nsukka / International Journal of Research in Arts and Social Sciences, 4(1), 120-135. https://academicexcellencesociety.com/ |
[58]
, p. 23); or: “analysis and synthesis are the complementary parts of a combined regressive-progressive method” (
, p. 15). Therefore, when Conklin (1908), Morgan (1917/1926), Dixon (1936), and others only analyzed the behaviors of cells into the reductionistic law of genetic mechanism, they had simply completed half of the task; their biology lacked a higher synthesis and remained basically 1-dimensional. As Fontaine insightfully pointed out, Just had added a great deal to this project in such theoretical statements describing his effort to find “a possible starting-point from which we may begin an attempt at the union, nowadays seemingly hopeless, of genetics and the physiology of development” (
, p. 70); or that “the physiology of development and heredity are merely two aspects of the life-history” (
| [43] | Just, E. E. (1936) A Single Theory for the Physiology of Development and Genetics. The American Naturalist, 70(728), 267-312. https://doi.org/10.1086/280666 |
[43]
, p. 272); or that “though we resolve [the ova] into constituent parts the better to understand it, we hold it as an integrated thing, as a unified system” (
| [45] | Just, E. E. (1939a) The Biology of the Cell Surface. P. Blakiston, Philadelphia. |
[45]
, p. 369).
It is interesting to note at this juncture, moreover, that Kant’s analytic-synthetic method has found new extensions within 20
th century philosophy. If an analyzed phenomenon has been demonstrated to be related to another analyzed phenomenon, it might have conceivably been found to have been related to a 3
rd, 4
th, or 5
th phenomenon, in an alternative synthesis of phenomena or even an underlying law. If an analyzed law has been demonstrated to be related to another analyzed law, it might conceivably have been found to be have been related to a 3
rd, 4
th or 5
th law, in an alternative synthesis of laws or even an underlying idea. The neo-Kantian philosopher Ernst Cassirer (1874-1945) had outlined this possibility of a “poly-synthetic method” in his books
Substance and Function (1910) and
Philosophy of Symbolic Forms (1923/1925). He based the possibility of his neo-Kantian theory on the poly-synthetic languages of Native Amerindians and others where subject and object effect the verb structure differently (
| [9] | Cassirer, E. A. (1923/2020) The Philosophy of Symbolic Forms, v.1. Yale Press, New Haven. |
[9]
, p. 276). However, while “with the help of the concept of relation, Cassirer justified the plurality of synthesizing acts, [b]y the term ‘plurality,’ Cassirer did not mean worlds with different natural laws but different modes of existence in one world” (
| [49] | Katsur, I. (2023) From Monism to Pluralism: Cassirer’s Interpretation of Kant. Вестник Российского университета дружбы народов. Серия: Философия, 27(3), 556-567. https://philpapers.org/rec/KATFMT-2 |
[49]
, p. 559). Nonetheless, it is an interesting and largely unassessed idea in biological sciences. Just’s “attempt at the union, […] seemingly hopeless, of genetics and the physiology of development” (
, p. 70) was certainly an important first step in advancing this analytic-synthetic idea, and it is significantly founded upon Kant’s
Critique of Pure Reason and its forgotten methodologies.
(2) By Fontaine’s designation of Just as the scientist “who has most closely approximated an artificial creation of life” (
| [20] | Fontaine, W. T. (1939) Philosophical Implications of the Biology of Dr. Ernest E. Just. The Journal of Negro History, 24(3), 281-290. https://doi.org/10.2307/2714381 |
[20]
, p. 281), a consequential biological question with a history traceable to at least the late 18
th century, had been imputed of relevance to his experiments. Because of German language translation questions, it is even possible that Kant had directly addressed some of these distinctions between: normal development (as it varied from species to species), parthenogenesis (organic development in a species without fertilization), and abiogenesis (creation of life from non-living matter). One scholarly evaluation of these translations by Ubirajara Azevedo-Marques has noted: “[I]f the adoption of spontaneous generation to translate Selbstgebärung [...] would have in its favor the supposed philological-theoretical adjust to the original expression and the relative proximity between this and the generatio æquivoca of seventy pages later, parthenogenesis [...] would fit better, allusively, either the explanation for the immediately following text («without impregnation by experience»), or the preformist vocabulary present therein” (
| [3] | Azevedo-Marques, U. (2015) On Epigenesis: Historical and Philological Remarks. In: Kant’s Lectures/ Kants Vorlesungen, Eds. B. Dörflinger, C. LaRocca, & R. Louden. De Gruyter, Berlin, pp. 261-272. https://doi.org/10.1515/9783110351538 |
[3]
, p. 265). The experimental basis for Kant’s statement was likely have been those of Charles Bonnet (1720-1793), also known as the first writer to have applied the term ‘evolution’ to a biological context. Additionally, for Kant’s texts, there is an “option to translate Selbstgebärung into English would be «self-generation» – an expression, by the way, already employed in the seventeenth century” (
| [3] | Azevedo-Marques, U. (2015) On Epigenesis: Historical and Philological Remarks. In: Kant’s Lectures/ Kants Vorlesungen, Eds. B. Dörflinger, C. LaRocca, & R. Louden. De Gruyter, Berlin, pp. 261-272. https://doi.org/10.1515/9783110351538 |
[3]
, pp. 265-266). In the
Critique of Judgment, Kant had upheld a cautious skepticism about the theoretical possibility of spontaneous generation (generatio æquivoca) or abiogenesis (Selbstgebärung). The following passages demonstrate both the expansive nature of Kant’s thought on the subject as well as his philosophical propriety in not making assumptions beyond what can be known by experiments:
“For the different animal genera approach one another gradually: from the genus where the principle of purposes seems to be bourne out most, namely, man, all the way to the polyp, and from it even to mosses and lichens and finally to the lowest stage of matter discernable to us, crude matter. From this matter, and its forces governed by mechanical laws (like those it follows in crystal formation), seems to stem all the technic that nature displays in organized beings […] This kind of generation is not absurd, as in
generatio æquivoca, which is the production of an organized being by the mechanics of crude, unorganized matter. Rather, this generation would still be a
generatio univoca in the most general sense of the world, because anything organic would be produced only from something else that is also organic, even though different in kind from it among beings of that type, as when, e.g., certain aquatic animals developed gradually into marsh animals and from these, after several generations, into land animals. This is not inconsistent
a priori, in the judgment of mere reason. Experience however does not show an example of it.” (
| [48] | Kant, I. (1790 [1987]) Critique of Judgment. Trans & Ed. W. S. Pluhar. Hackett Press, Indianapolis. |
[48]
, pp. 304-305).
Similarly, Just outlined a need for hesitancy in concluding that abiogenesis had been achieved or if it was even possible. Indeed he had stated that “we do not know that there is no discontinuity between the non-living and the living world” (
| [45] | Just, E. E. (1939a) The Biology of the Cell Surface. P. Blakiston, Philadelphia. |
[45]
, p. 12) and that the matter remained unsettled, indeed with regard to certain philosophical considerations. Moreover, as has been noted, Just’s experimental research had found that “the conditions for successful artificial initiation of development are more exacting” (
| [35] | Just, E. E. (1915) An Experimental Analysis of Fertilization in Platynereis Megalops. The Biological Bulletin, 28 (2), 93-114. https://doi.org/10.2307/1536272 |
[35]
, p. 109) than what had been commonly supposed by other biologists.
However, it is also possible to read within Just’s published works that he had actually made abstract theoretizations far beyond these loosely stated Kantian-type restrictions upon abiogenetic experiments. After having studied in detail the process of blastomere pattern formation and the role of cytoplasmic potencies both controlling and in turn being reciprocally restricted by the nucleus, Just perceived the essential nature of the whole-part relation in biological organization. This is what he added into the discussed question of the origin of life. In Kant’s defense, he had similarly discussed how in development organisms “combine into the unity of a whole because they are reciprocally cause and effect of their form” (
| [48] | Kant, I. (1790 [1987]) Critique of Judgment. Trans & Ed. W. S. Pluhar. Hackett Press, Indianapolis. |
[48]
, p. 252). However, it does not seem that Kant further applied this whole-part metaphysical relation to the first evolutionary emergence of life from a non-living state. Ernest Everett Just extended this whole-part relation to this very question:
“Even if we assume that the primordial living thing was a mass of homogeneous protoplasm structurally the same throughout, there must have early arisen a differentiation between surface and interior. In the constant interchange between environment and organism reactions must have taken place first in the more superficially located cytoplasmic structure; these reactions would condition succeeding ones in the endoplasm. The first step in the evolutionary process, then, was a differentiation of the cytoplasm into ectoplasm and endoplasm. The second step, according to this theory, was a nucleo-cytoplasmic differentiation. We have thus a picture of the primordial living thing as a mass composed of the prototype of the ground-substance in cells as we know them to-day which limited itself in space by a changed surface, its ectoplasm” (
| [45] | Just, E. E. (1939a) The Biology of the Cell Surface. P. Blakiston, Philadelphia. |
[45]
, p. 357).
This broad view of early evolution given by Just might have provided a key insight into the possibility of a neo-Kantian philosophy of abiogenetic evolution, specifically, by extending the whole-part relation to the primordial environment and not just the tissues-organs/whole-part of a living form. For its full theoretical development, it is definitely not clear that this approach has been yet attempted by 20
th century mathematical biologists. Some researchers do occasionally invoke Kant for the whole-part relation
, and experiments like those of Miller-Urey have demonstrated the role of the environment in biochemical reactions necessary for emergence of life
. However, it does not seem that the full complex-mathematical description of this dynamic process, viz., whole (environment) – part (organism) — whole (organism) – part (biochemical constituent), etc., etc., has been devised in terms of field equations. Goodwin, for example, had considered Kant’s theory as far as organisms as “self-generating wholes, complex systems that obey principles of dynamic order described by field equations [and] reproducing structures of a distinct kind whose transformations defined the range of living forms available for evolution” (
, pp. 53-54). But his work had presupposed structures that already had the ability to reproduce, without considering the initially homogeneous primordial state between the interior and exterior of living matter, and thus left unanswered more fundamental questions in evolutionary theory.
(3) Fontaine’s paper on Just, arguably, has charted a profoundly revolutionary path forward for these loose ends clearly found in earlier mathematical biology. Based on his suggested parallel to the text Space, Time and Deity (1920) by Samuel Alexander, it is accordingly asked: are higher dimensions and alternative geometries applicable in the field of theoretical biological? With a background in philosophy of mathematics and philosophy of psychology from Oxford and Freiburg universities, Alexander’s 1916-1918 Gifford Lectures (published in 1920) were given among those of other lecturers invited to speak for multiple, successive years. His hundreds of pages of ideas presented therein provide rich source material for the applicability and meaning of higher-dimensional geometries:
“There are many geometries though there is but one space. Strictly speaking, it is only by a mistake of language that we speak of non-Euclidean space or even of Euclidean space; we have only Euclidean or non-Euclidean geometry [...] This [...] applies to the non-Euclidean geometries of Lobatchewsky and others, the so-called hyperbolic and elliptic geometries. Now, in the case of these geometries, the question does not arise whether they take us into a world different from our experienced space. They are merely different systems of explaining, not the ultimate nature of space, but its behaviour in detail. They employ different postulates. At the same time they introduce us to another feature of geometry and of mathematics generally, its method of generalisation. Euclidean geometry is only one instance of geometry of empirical space” (
| [1] | Alexander, S. (1920a) Space, Time, and Deity: Gifford Lectures, v.1. Macmillan, London. |
[1]
, p. 157).
By distinguishing between space as a geometric construction and space as an empirical perception, Alexander had indirectly followed the Kantian paradigm of distinguishing between space as a pure form of intuition and space as an empirical reality, however, which for Kant, were unified into an absolute space without any possible points of nexus for non-Euclidean or higher-dimensional space to bear upon either, as Kauark-Leite has shown
| [50] | Kauark-Leite, P. (2017) On the Epistemic Status of Absolute Space: Kant’s Directions in Space Read from the Standpoint of his Critical Period. Kant-Studien, 108(2), 175-194. https://doi.org/10.1515/kant-2017-0015 |
[50]
. Alexander, elsewhere in the text, compared these higher or non-Euclidean geometries to “scaffoldings” that have “their utility in their application” as well as “a value irrespective of utility” (
| [1] | Alexander, S. (1920a) Space, Time, and Deity: Gifford Lectures, v.1. Macmillan, London. |
[1]
, p. 162).
While Alexander had ventured to bring higher-geometries into a philosophical basis for application to empirical scientific investigations, it might have been proscribed by him as to its detailed applicability in evolutionary biology. E.g. “since time is infinite it might seem that every form of existence must have existed in the past […], and therefore, in the strictest sense, the universe is not an evolution at all, but the whole of its varied riches exists already no matter at what point in the history we are imagined to stop” (
| [2] | Alexander, S. (1920b) Space, Time, and Deity: Gifford Lectures, v.2. Macmillan, London. |
[2]
, p. 336). In other words, Alexander’s utilitarian applications of higher-dimensional or non-Euclidean geometries to the questions of the origin of life, at least in this section of the text, were blurred out in the infinity of possibilities. However, other statements in
Space, Time and Deity left open these questions, perhaps of vital importance for a mathematical study of biology:
“As we ascend the scale of being in the order of time aggregates are replaced by organic systems; and the higher a thing is in the scale, the greater it seems is its ordered complexity. But system in general exists in every complex even in the least organised, all disorder has its own complex plan [...] The nature of an organism and still more a work of art is rightly exemplary in the methods which reason follows. Thought, in following the clue of coherence amongst its data, as science always does, is thus bringing back the scattered members of the universe into the spatio-temporal continuity out of which, in spite of their disguises of qualities higher than mere motion, they ultimately sprang” (
| [1] | Alexander, S. (1920a) Space, Time, and Deity: Gifford Lectures, v.1. Macmillan, London. |
[1]
, p. 237).
Figure 2. (L) William Fontaine from University of Pennsylvania’s Archives and Records; (R) Book cover of Humanities Press edition of Samuel Alexander’s Space, Time and Deity (1920).
The publications of Just throughout the 1920s and 1930s, as well as Fontaine’s insightful 1939 article on this work, followed chronologically closely upon these advances in higher-dimensional mathematics and Alexander’s philosophical suggestions of their utility of application in these fields. Certainly, these three distinct scholarly fields, viz., biology, mathematics and philosophy, which Fontaine had endeavored to bring together, have led to many fruitful developments in the 20th-century. Yet more studies relating all of these subjects are still nonetheless wanting to this day.
4. Neo-Kantian Discussion of Higher-Dimensional Unified Theories
Kant’s fundamental commitments to 3-dimensional space are seen by most philosophers to be characteristic of his entire corpus of texts. For example, Gardner (1969) has noted that his consideration of incongruent left/right objects led him by the time of his publication of the
Critique of Pure Reason (1781/1787) to argue that “there must be some basis, some ground for calling it ‘left’ even when it is the sole object in the universe. Kant could see no way of providing such a ground except by assuming that space itself possessed some sort of absolute, objective structure — a kind of 3-dimensional lattice that could furnish a means of defining the handedness of a solitary asymmetric object” (
| [22] | Gardner, M. (1969) The Ambidextrous Universe: Left, Right, and the Fall of Parity. Signet Books, Boston. |
[22]
, p. 147). He noted further that “a modern reader familiar with
n-dimensional geometry should have little trouble seeing through the verbal confusion of Kant's thought experiment” (
| [22] | Gardner, M. (1969) The Ambidextrous Universe: Left, Right, and the Fall of Parity. Signet Books, Boston. |
[22]
, p. 147). Similarly, Van Cleve (1987) has noted that “in 1768, Kant believed that the existence of such objects furnished proof of a Newtonian or absolutist as against a Leibnizian or relationist view of the reality of space: space is a thing in its own right, not just a construction out of material bodies and the relations among them” (
, p. 33), because it is clear that in a “4-dimensional space, a right hand could be flipped over so as to become its own incongruent counterpart.” (
, p. 44). Unique among those who follow Kant’s geometric ideas is Friedman (1992). He has pointed out that Kant had speculated in a 1747 text “that there may be other actual worlds (not in connection with our world) that have spaces of more than 3-dimensions” (
| [21] | Friedman, M. (1992) Kant and the Exact Sciences. Harvard Press, Boston. |
[21]
, p. 33), and that Kant’s statement on the “impossibility of our imagining a space of other than 3-dimensions is equally contingent and depends simply on [...] the very same arbitrary law [i.e. God's original choice]” (
| [21] | Friedman, M. (1992) Kant and the Exact Sciences. Harvard Press, Boston. |
[21]
, p. 27). Moreover, Friedmann has found a loophole in other statements by Kant against higher-dimensional space, an original argument against absolute space that Kant himself had first posited: “it is precisely by replacing Leibniz’s conception of the ideality of space with his own conception of the fundamentally dynamical character of space that Kant himself has first exposed geometry to the threat of empirical disconfirmation” (
| [21] | Friedman, M. (1992) Kant and the Exact Sciences. Harvard Press, Boston. |
[21]
, p. 27).
As has been noted, Just and Loeb had integrated the language of mathematical physics into their biological philosophy. When Just wrote in 1936 on the theory of genetic-mechanism that “at
critical points the theory breaks down. Because it so frequently extends itself too far into the realm of the unknown, it makes too great demands on credulity” (
| [43] | Just, E. E. (1936) A Single Theory for the Physiology of Development and Genetics. The American Naturalist, 70(728), 267-312. https://doi.org/10.1086/280666 |
[43]
, p. 291)— he was echoing statements like those of Einstein in 1921:— “Euclidean metrics can no longer be valid with respect to accelerated systems of co-ordinates [...] this enormous difficulty was mitigated by our knowledge that Euclidean metrics [still] hold for small domains” (
, p. 784). Others during that time period used similar terminology for such innovative quests to enter into unknown space-time patterns; for example, Dorothy Wrinch (1894-1976) wrote thus in 1922:
[…] is one of a very large number of forms which satisfy the Einstein conditions. [Paul] Painlevé [(1863-1933)] cites some of the other possible forms for the relation between the length element and the four co-ordinates (
), and indicates the various consequences which ensue according to the particular form adopted [...] [I]t is evident that there are at least as many functions satisfying these conditions as there are points in the mathematical continuum. This difficulty of choosing between a set of functions all of which satisfy the data of the problem presents itself at several
critical points of the Einstein theory” (
, p. 381).
Nonetheless, there were additional writers, for example Spaulding (1918), who characterized the critical points of transitioning between two alternative theories as being a philosophical task, perhaps along the lines of the poly-synthetic of Cassirer:
“[T]he close resemblance between the situations in geometrical science and in philosophy [is] not complete. The several geometries, e.g. the Euclidean, the Lobatchewskian, and the Riemannian, are each self consistent and free from internal contradiction. This, however, cannot be said of the several philosophical systems some of which are very evidently self-contradictory and self-refuting at
critical points [...] Bolyai in 1832 and Lobatchewsky in 1835 independently found it possible to develop a consistent system of geometry by denying the Euclidean axiom of parallels [...] The non-Euclidean system that results is self consistent and quite as applicable to our space as the traditional Euclidean geometry, as far as empirical measurement can determine” (
| [69] | Spaulding, E. G. (1918) The New Rationalism: The Development of a Constructive Realism upon the Basis of Modern Logic and Science. H. Holt Press, New York. |
[69]
, p. 6).
In both cases, critical points were introduced to evaluate the ranges of validity of certain concepts. Theories, apparently, break down at certain points, and questions arise as to why such ranges of validity emerge and where more fundamental theories are to be formulated. Could the unification be in higher dimensional spaces?
This possibility has certainly gained traction recently in theoretical physics through the advancement of ideas associated with grand unified theories and string theories. Usually, particle physics tests theories by dividing subatomic matter into even smaller elements (baryons, leptons, photons, etc.), and there is no immediate relevance to large biological systems, where these particles are assembled into molecules, organelles, cells, and tissues. However, there might be new theoretical discoveries that would challenge this division of the sciences. A recent paper by Hong Wang, Xinyu Li and Jin Wang (2021) has described the origin of thermodynamic irreversibility in a 5-dimensional space, and certain biologically relevant concepts, viz: chemical evolution, lysis, cell fertilization, tissue differentiation, aging, etc., are all irreversible processes. The essence of their theory can be stated succinctly:
"4-dimensional Euclidean
universe could be viewed as a 5-dimensional sphere’s surface in higher dimensional Euclidean space [...] If all the universes are
vacuum, the detailed balance is preserved [...] Recent studies show that there is a possibility of the tunneling transition from Minkowski or
vacuum to
universes through the bounce […] [I]f we choose suitable boundary conditions, the detailed balance can be broken down [...] The flux associated with the detailed balance breaking provides the dynamical origin of the irreversibility and time arrow of the multiverse” (
| [74] | Wang, H., Li, X., Wang, J. (2021) Quantifying the Potential and Flux Landscapes for Nonequilibrium Multiverse, A New Scenario for Time Arrow. Journal of High Energy Physics, 2(1), 1-23. https://doi.org/10.1007/JHEP02(2021)105 |
[74]
, p. 6).
Thus, the elusive concept of thermodynamic irreversibility, which is perhaps impossible to imagine in 3-dimensions, and which perplexes the mind when the 4th-dimension bears no similarity to the spatial-dimensions, has found a surprising theoretical justification in 5-dimensions and Minkowski spaces. Perhaps the contributions of higher-dimensional mathematics are indeed relevant to the biological sciences through the emergence of thermodynamic irreversibility.
Other metaphysical descriptions of these extra dimensions tend to be provided by generalists and would require further interpretation for determining their biological relevance. As Rieger (2002) has written: “a physicist will adjust the physics, not the mathematics, whereas a biologist will adjust the biology, not the physics. From a logical point of view, however, the global theory is up for revision too, and in drastic enough circumstances will be abandoned” (
, p. 249). Thus biological theories in general do not tend to shape the future direction of physical theories toward higher dimensions, but conversley. Considered along these lines, for example, Williams (2014) has summarized the higher dimensions of superstring theory without any direct interpretation in biological theories: the 1
st-dimension is length in x-axis; the 2
nd-dimension adds height in the y-axis; the 3
rd-dimension adds depth in the z-axis; the 4
th-dimension adds time which relates matter to other matter as it changes along temporal points; the 5
th-dimension adds a means of measuring distances between our world and other similar worlds; the 6
th-dimension maps this into a plane where you could compare all possible worlds that start with the same initial conditions (and travel forward and backward through them); in the 7
th-dimension is added a measurement between possible worlds that start with different initial conditions; in the 8
th-dimension is added the plane of possible universes with different initial conditions; in the 9
th-dimension it becomes possible to measure distances between all possible universe with different laws of physics and different initial conditions; finally, in the 10
th-dimension there is a representation of every possible set of laws, every set of initial conditions, and every path of particular trajectories of all universes envisioned in a complete whole
. These higher dimensions are described mathematically by a Calabi-Yau manifold, a topic of very active research in mathematics and theoretical physics
| [28] | Greene, B. (2003) The Elegant Universe: Superstrings, Hidden Dimensions, and the Quest for the Ultimate Theory. Norton, London. |
| [75] | Williams, M. (2014) A Universe of 10 dimensions. PhysOrg https://phys.org/news/2014-12-universe-dimensions.html Accessed 24 November 2024. |
[28, 75]
, however, without much direct bearing on biological theories. In the other direction of this trend, however, are recent papers by Birch et al. (2020)
and Dung (2024)
that have provided a 5-dimensional analysis of animal and human consciousness
. How these might map (isomorphic, homomorphic, heteromorphic) to the spatial dimensions of superstring theory remain to be explored.
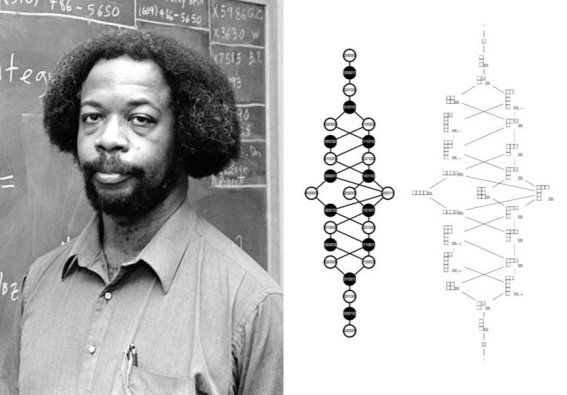
Figure 3. (L) Sylvester James Gates photo from American Institute of Physics, Emilio Segrè Visual Archives; (R) Adinkras (i.e. supersymmetric algebras) of 10- or 11-dimensions, depicted in Gates, Hu & Mak (2021), can be used to predict the structure of gravitons, in a way analogous to genetic codons’ predictions of protein structure
Another recent contribution to the topic of higher-dimensional mathematics and its applications in biological systems has been published by Sylvester James Gates. Using 10-dimensional and 11-dimensional algebraic sets, he and collaborators have made predictions about the possible structures of gravitons associated with each particular set — analogously, as he has noted, genes and the 3-letter codons can be used to predict protein structures. It is indeed conceivable, therefore, that biological structures involving whole-part relations could be based upon predictions in these higher-dimensional geometries; however, Gates et al.’s research has already taken us in the first step in that direction:
“By the study of genes and knowing their expressions, one can deduce information about structures. This is the reason why using the foundation of the adinkra concept, we were able to analyze all the 2
31 (= 2,147,483,648) bosonic degrees of freedom and all the 2
31 (= 2,147,483,648) fermionic degrees of freedom in the 11D, N = 1 scalar superfield. Scalar superfields act as gateways to the similar deciphering the component eld spectra of superfields in all spin representations. Using this fact, we have begun the task of identifying superfields that contain the conformal graviton in these contexts” (
| [25] | Gates, S. J., Hu, Y., Mak, S. (2021) Advening to Adynkrafields: Young Tableaux to Component Fields of the 10D, N= 1 Scalar Superfield. Advances in Theoretical and Mathematical Physics, 25(1), 1449-1547. https://doi.org/10.4310/ATMP.2021.v25.n6.a3 |
[25]
, p. 4).
Working with his co-authors Yangrui Hu and S-N. Hazel Mak, Gates has been able to give a presentation of his research at the National Institute of Health on 26 April 2022, with a talk that was entitled: “How Looking at Genetics and Networks Led to Solving a Quantum Gravity Problem” (https://www.youtube.com/watch?v=TdhecXVbX5o). It is also interesting and valuable to note that Gates advocates for minority representation in the sciences as well as the essential element of creativity when developing scientific theories
| [23] | Gates, S. J. (2012) Science, Irrationality & Innovation. In: Scientific Work and Creativity: Advice from the Masters, Eds. R. Smith, Citizen Scientists League Press, Clearwater FL, pp 119-139. |
| [24] | Gates, S. J. (2023) Expelled from the Mountain Top? Science, 380 (6649), 993-993. https://doi.org/10.1126/science.adj0265 |
[23, 24]
.