The study evaluated response to selection from within-breed selection strategy for conventional (CBS) and genomic (GBS) breeding schemes. These breeding schemes were evaluated in both high-health environments (nucleus) and smallholder farms (commercial). Breeding goal was to develop a dual-purpose IC for both eggs and meat through selective breeding. Breeding objectives were body weight (BW), egg weight (EW), egg number (EN) and resistance to Newcastle disease (AbR). A deterministic simulation was performed to evaluate rates of genetic gain and inbreeding. Base population in the nucleus was made up of 40 cockerels and 200 pullets. Selection pressure was 4% and 20% in the males and the females, respectively. The impact of nucleus size and selection pressure on rates of genetic gain and inbreeding of the breeding program was investigated through sensitivity analysis. SelAction software was used to predict rates of genetic gain and inbreeding. Results showed that using CBS in the nucleus, the breeding goal was 340.41$ and 1.13 times higher than that in the commercial flock. Inbreeding rate per generation of selected chicken in the nucleus was 1.45% and lower by 1.32 times compared to their offspring under smallholder farms. Genetic gains per generation in the nucleus for BW and EN traits were 141.10 g and 1.07 eggs and 1.12 and 1.10 times greater than those in smallholder farms, respectively. With GBS, the breeding goal was increased by 3.00 times whereas inbreeding rate was reduced by 3.15 times. Besides, using GBS, the selected birds in the nucleus were relatively similar to those in a commercial environment. Finally, the study revealed that the nucleus size and mating ratio influence the rates of genetic gain and inbreeding in both GBS and CBS. This study shows that IC in Rwanda have the potential to be improved through within-breed selection strategy using either CBS or GBS.
| Published in | Animal and Veterinary Sciences (Volume 12, Issue 3) |
| DOI | 10.11648/j.avs.20241203.13 |
| Page(s) | 95-106 |
| Creative Commons |
This is an Open Access article, distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution and reproduction in any medium or format, provided the original work is properly cited. |
| Copyright |
Copyright © The Author(s), 2024. Published by Science Publishing Group |
Genetic Gain, Inbreeding, Indigenous Chicken, Selection, Rwanda
Traits | Correlations | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
BW | EN | EW | AbR | ENC | BWC | BWGS | ENGS | EWGS | AbRGS | BWCGS | ENCGS | |
Var (P) | 139,929.56 | 130.69 | 18.80 | 5,677,315.41 | ||||||||
EV (US$) | 2.15 | 0.19 | -0.001 | -0.23 | ||||||||
BW | 0.24 | 0.22 | 0.10 | -0.07 | 0.17 | 0.75 | 0.34 | 0.06 | 0.04 | -0.03 | 0.26 | 0.04 |
EN | 0.23 | 0.24 | -0.19 | -0.04 | 0.76 | 0.17 | 0.08 | 0.25 | -0.08 | -0.01 | 0.06 | 0.19 |
EW | 0.20 | -0.13 | 0.44 | -0.01 | -0.14 | 0.15 | 0.03 | -0.04 | 0.44 | -0.00 | 0.05 | -0.03 |
AbR | -0.07 | -0.02 | -0.00 | 0.27 | -0.03 | -0.05 | -0.02 | -0.01 | 0.00 | 0.36 | 0.02 | -0.00 |
ENC | 0.04 | 0.16 | -0.02 | -0.00 | 0.24 | 0.22 | 0.06 | 0.19 | -0.06 | -0.01 | 0.06 | 0.25 |
BWC | 0.18 | 0.04 | 0.04 | -0.01 | 0.23 | 0.24 | 0.26 | 0.04 | 0.07 | -0.02 | 0.26 | 0.06 |
BWGS | 0.17 | 0.07 | 0.03 | -0.02 | 0.06 | 0.25 | 0.97 | 0.03 | 0.02 | -0.01 | 0.14 | 0.02 |
ENGS | 0.05 | 0.09 | -0.04 | -0.01 | 0.19 | 0.04 | 0.03 | 0.97 | -0.03 | -0.00 | 0.02 | 0.08 |
EWGS | 0.04 | -0.08 | 0.29 | -0.00 | -0.06 | 0.07 | 0.02 | -0.03 | 0.97 | -0.00 | 0.04 | -0.02 |
AbRGS | -0.03 | -0.01 | -0.00 | 0.19 | -0.01 | -0.02 | -0.01 | -0.00 | -0.00 | 0.97 | -0.01 | -0.00 |
BWCGS | 0.25 | 0.06 | 0.05 | -0.02 | 0.08 | 0.25 | 0.14 | 0.02 | 0.04 | -.0.01 | 0.97 | 0.03 |
ENCGS | 0.04 | 0.19 | -0.02 | -0.00 | 0.11 | 0.05 | 0.02 | 0.07 | -0.02 | -0.00 | 0.03 | 0.97 |
Scheme | Response (US$) | Rate of inbreeding (%) | Accuracy of index | |
|---|---|---|---|---|
CBS | Nucleus | 340.41 | 1.45 | 0.55 |
Commercial | 301.17 | 1.91 | 0.47 | |
GBS | Nucleus | 1,024.45 | 0.46 | 0.97 |
Commercial | 1,024.44 | 0.46 | 0.97 |
Trait | CBS | GBS | ||
|---|---|---|---|---|
Nucleus | Commercial | Nucleus | Commercial | |
BW | 158.24 | 141.39 | 476.44 | 476.44 |
EN | 1.07 | 0.97 | 0.49 | 0.48 |
EW | 0.24 | 0.43 | 0.13 | 0.20 |
AbR | -82.10 | -72.17 | -45.39 | -30.37 |
| [1] | Adeleke, M. A., Peters, S. O., Ozoje, M. O., Ikeobi, C. O. N., Bamgbose, A. M., Adebambo, O. A., 2011. Growth performance of Nigerian local chickens in crosses involving an exotic broiler breeder. Trop. Anim. Health Prod. 43, 643–650. |
| [2] | Assan, N., 2015. Prospects for indigenous chickens genetic improvement and conservation in Zimbabwe. Agric. Adv. 4, 49–56. |
| [3] | Besbes, B., Gibson, J. P., 1999. Genetic variation of egg production traits in purebred and crossbred laying hens. Anim. Sci. 68, 433–439. |
| [4] | Bett, H. K., Peters, K. J., Bokelmann, W., 2011. Hedonic price analysis to guide in breeding and production of indigenous chicken in Kenya. Livest. Res. Rural Dev. 23, 142. |
| [5] | Bijma, P., 2000. Long-term Genetic Contributions - Prediction of Rates of Inbreeding and Genetic Gain in Selected Populations, Animal Breeding and Genetic Group, Department of Animal Sciences. |
| [6] | Bytyqi, H., Fuerst-Waltl, B., Mehmeti, H., Baumung, R., 2015. Economic values for production traits for different sheep breeds in Kosovo. Ital. J. Anim. Sci. 14, 603–609. |
| [7] | Cervantes, I., Gutiérrez, J. P., Meuwissen, T. H. E., 2016. Response to selection while maximizing genetic variance in small populations. Genet. Sel. Evol. 48, 69. |
| [8] | Chen, J., Wang, Y. chun, Zhang, Yi, Sun, D. xiao, Zhang, S. li, Zhang, Yuan, 2011. Evaluation of breeding programs combining genomic information in chinese holstein. Agric. Sci. China 10, 1949–1957. |
| [9] | Christensen, O. F., Legarra, A., Lund, M. S., Su, G., 2015. Genetic evaluation for three-way crossbreeding. Genet. Sel. Evol. 47, 1–13. |
| [10] | Chu, T. T., Bastiaansen, J. W. M., Berg, P., Romé, H., Marois, D., Henshall, J., 2019. Use of genomic information to exploit genotype ‑ by ‑ environment interactions for body weight of broiler chicken in bio ‑ secure and production environments. Genet. Sel. Evol. 1–13. |
| [11] | Daetwyler, H. D., Calus, M. P. L., Pong-Wong, R., de los Campos, G., Hickey, J. M., 2013. Genomic prediction in animals and plants: Simulation of data, validation, reporting, and benchmarking. Genetics 193, 347–365. |
| [12] | Daetwyler, H. D., Villanueva, B., Bijma, P., Woolliams, J. A., 2007. Inbreeding in genome-wide selection. J. Anim. Breed. Genet 124, 369–376. |
| [13] | Daetwyler, H. D., Villanueva, B., Woolliams, J. A., 2008. Accuracy of predicting the genetic risk of disease using a genome-wide approach. PLoS One 3. |
| [14] | Dekkers, J. C. M., 2007a. Prediction of response to marker-assisted and genomic selection using selection index theory. J. Anim. Breed. Genet. 124, 331–341. |
| [15] | Dekkers, J. C. M., 2007b. Marker-assisted selection for commercial crossbred performance. J. Anim. Sci. 85, 2104–2114. |
| [16] | Dekkers, J. C. M., Chakraborty, R., 2004. Optimizing purebred selection for crossbred performance using QTL with different degrees of dominance. Genet. Sel. Evol. 36, 297–324. |
| [17] | Duenk, P., Calus, M. P. L., Wientjes, Y. C. J., Breen, V. P., Henshall, J. M., Hawken, R., Bijma, P., 2019. Estimating the purebred-crossbred genetic correlation of body weight in broiler chickens with pedigree or genomic relationships. Genet. Sel. Evol. 51, 1–11. |
| [18] | Ebrahimpourtaher, S., Alijani, S., Rafat, S. A., Sharifi, A. R., 2018. Potential of Genomic Breeding Program in Iranian Native Chickens. Iran. J. Appl. Anim. Sci. 8, 305–315. |
| [19] | Eggen, A., 2012. The development and application of genomic selection as a new breeding paradigm. Anim. Front. 2, 10–15. |
| [20] | Esfandyari, H., Berg, P., Sørensen, A. C., 2018. Balanced selection on purebred and crossbred performance increases gain in crossbreds. Genet. Sel. Evol. 50, 1–14. |
| [21] | Esfandyari, H., Sørensen, A. C., Bijma, P., 2015. Maximizing crossbred performance through purebred genomic selection. Genet. Sel. Evol. 47, 1–16. |
| [22] | Falconer, D. S. (Douglas S., Mackay, T. F. C., 1996. Introduction to quantitative genetics. Longman. |
| [23] | Groenen, M. A. M., Wahlberg, P., Foglio, M., Cheng, H. H., Megens, H. J., Crooijmans, R. P. M. A., Besnier, F., Lathrop, M., Muir, W. M., Wong, G. K. S., Gut, I., Andersson, L., 2009. A high-density SNP-based linkage map of the chicken genome reveals sequence features correlated with recombination rate. Genome Res. 19, 510–519. |
| [24] | Gwaza, D. S., Dim, N. I., Momoh, O. M., 2016. Genetic improvement of egg production traits by direct and indirect selection of egg traits in Nigerian local chickens. Adv. Genet. Eng. 05. |
| [25] | Habimana, R., Okeno, T. O., Ngeno, K., Mboumba, S., Assami, P., Keambou, C. T., Breeding, A., Group, G., Eastern, B., 2018. Four gene pools of Rwandese indigenous chicken based on microsatellite markers, in: Proceedings of the World Congress on Genetics Applied to Livestock Production, 11.874. pp. 1–4. |
| [26] | Hazel, L. N., 1943. The genetic basis for constructing selection indexes. Genetics 28, 476–90. |
| [27] | Hennnatabadi, R. N., Seidavi, A., Gharahveysi, S., 2011. Estimation response to selection in three commercial lines of silkworm (Bombyx mori L.) having peanut cocoon. J. Anim. Vet. Adv. 10, 2208–2214. |
| [28] | Henryon, M., Berg, P., Sørensen, A. C., 2014. Invited review: Animal-breeding schemes using genomic information need breeding plans designed to maximise long-term genetic gains. Livest. Sci. 166, 38–47. |
| [29] | Janssen, K., Saatkamp, H. W., Calus, M. P. L., Komen, H., 2019. Economic optimization of full-sib test group size and genotyping effort in a breeding program for Atlantic salmon. Genet. Sel. Evol. 51, 1–14. |
| [30] | Kariuki, C. M., Komen, H., Kahi, A. K., van Arendonk, J. A. M., 2014. Optimizing the design of small-sized nucleus breeding programs for dairy cattle with minimal performance recording. J. Dairy Sci. 97, 7963–7974. |
| [31] | Kaufmann, F., Das, G., Preisinger, R., Schmutz, M., König, S., Gauly, M., 2011. Veterinary Parasitology Genetic resistance to natural helminth infections in two chicken layer lines 176, 250–257. |
| [32] | Lamont, S. J., Pinard-van der Laan, M. H., Cahaner, A., Poel, J. J. van der, Parmentier, H. K., 2009. Selection for disease resistance: direct selection on the immune response., in: Poultry Genetics, Breeding and Biotechnology. pp. 399–418. |
| [33] | Leitch, H. W., Smith, C., Burnside, E. B., Quinton, M., 1994. Genetic Response and Inbreeding with Different Selection Methods and Mating Designs for Nucleus Breeding Programs of Dairy Cattle. J. Dairy Sci. 77, 1702–1718. |
| [34] | Liu, H., Henryon, M., Sørensen, A. C., 2017. Mating strategies with genomic information reduce rates of inbreeding in animal breeding schemes without compromising genetic gain. Animal 11, 547–555. |
| [35] | Liu, T., Qu, H., Luo, C., Li, X., Shu, D., Lund, M. S., Su, G., 2014. Genomic selection for the improvement of antibody response to newcastle disease and avian influenza virus in chickens. PLoS One 9, 1–6. |
| [36] | Lwelamira, J., 2012. Genotype-environmental (GxE) interaction for body weight for kuchi chicken ecotype of tanzania reared on-station and on-farm. Int. J. Poult. Sci. 11, 96–102. |
| [37] | Lwelamira, J., Kifaro, G. C., 2010. Desired-gain selection indices for improving performance of two Tanzania local chicken ecotypes under intensive management. African J. Agric. Res. |
| [38] | Magothe, T. M., Okeno, T. O., Muhuyi, W. B., Kahi, A. K., 2012. Indigenous chicken production in Kenya: I. Current status. Worlds. Poult. Sci. J. 68, 119–132. |
| [39] | Mahoro, J., 2017. Characterisation of production systems and development of breeding objectives for indigenous chicken in Rwanda. MSc. Thesis, Egerton University. |
| [40] | Mahoro, J., Muasya, T. K., Mbuza, F., Habimana, R., Kahi, A. K., 2017. Characterization of indigenous chicken production systems in Rwanda. Poult. Sci. 96, 4245–4252. |
| [41] | Mahoro, J, Muasya, T. K., Mbuza, F., Mbuthia, J., Kahi, A. K., 2017b. Farmers’ breeding practices and traits of economic importance for indigenous chicken in RWANDA. Trop. Anim. Health Prod. 50, 121–128. |
| [42] | Marcos-atxutegi, C., Gandolfi, B., Arangu, T., 2009. Veterinary Parasitology Antibody and inflammatory responses in laying hens with experimental primary infections of Ascaridia galli 161, 69–75. |
| [43] | Mbuthia, J. M., Rewe, T. O., Kahi, A. K., 2015. Analysis of pig breeding management and trait preferences in smallholder production systems in Kenya. Anim. Genet. Resour. génétiques Anim. genéticos Anim. 56, 111–117. |
| [44] | Mbuza, F., Denis, M., Janvier, M., Xavier, R., 2016. Characterization of low cost village Poultry production in Rwanda. Int. J. Livest. Prod. 7, 76–82. |
| [45] | Meuwissen, T., Hayes, B., Goddard, M., 2016. Genomic selection: A paradigm shift in animal breeding. Anim. Front. 6, 6–14. |
| [46] | Meuwissen, T. H. E., 1997. Managing inbreeding in selection and genetic conservation schemes of livestock. |
| [47] | Mohan, J., Sharma, S. K., Kolluri, G., Dhama, K., 2018. History of artificial insemination in poultry, its components and significance. Worlds. Poult. Sci. J. 74, 475–488. |
| [48] | Mtileni, B. J., Muchadeyi, F. C., Maiwashe, A., Chimonyo, M., Dzama, K., 2012. Conservation and utilisation of indigenous chicken genetic resources in Southern Africa. Worlds. Poult. Sci. J. 68, 727–748. |
| [49] | Muir, W. M., Cheng, H., Croney, C., 2014. Methods to address poultry robustness and welfare issues through breeding and associated ethical considerations 5, 1–11. |
| [50] | Mulder, H. A., 2016. Genomic selection improves response to selection in resilience by exploiting genotype by environment interactions. Front. Genet. 7, 1–11. |
| [51] | Mulder, H. A., Bijma, P., 2006. Benefits of cooperation between breeding programs in the presence of genotype by environment interaction. J. Dairy Sci. 89, 1727–1739. |
| [52] | Mulder, H. A., Bijma, P., 2005. Effects of genotype x environment interaction on genetic gain in breeding programs. J. Anim. Sci. 83, 49–61. |
| [53] | Mwambene, P. L., Kyallo, M., Machuka, E., Githae, D., Pelle, R., 2019. Genetic diversity of 10 indigenous chicken ecotypes from Southern Highlands of Tanzania based on Major Histocompatibility Complex-linked microsatellite LEI0258 marker typing. Poult. Sci. 98, 2734–2746. |
| [54] | Natukunda, K., Kugonza, D. R., Kyarisiima, C. C., 2011. Indigenous chickens of the Kamuli plains in Uganda: II. Factors affecting their marketing and profitability. Livest. Res. Rural Dev. 23. |
| [55] | Ndegwa, J. M., R. Mead, R., Norrish, P., Shephered, D. D., Kimani, C. W., Wachira, A. M., 2012. Growth Characteristics of Six Reciprocal Crosses of Kenyan Indigenous Chicken. J. Agric. Sci. 4. |
| [56] | Ndung’u, C. W., Okeno, T. O., Muasya, T. K., 2020. Pooled parameter estimates for traits of economic importance in indigenous chicken in the tropics. Livest. Sci. 1–34. |
| [57] | NISR, 2019. Rwanda Statistical YearBook. |
| [58] | Nwagu, B. I., Olorunju, S. A. S., Oni, O. O., Eduvie, L. O., Adeyinka, I. A., Sekoni, A. A., Abeke, F. O., 2007. Response of Egg Number to Selection in Rhode Island Chickens Selected for Part Period Egg Production. Int. J. Poult. Sci. 6, 18–22. |
| [59] | Okeno, T. O., Kahi, A. K., Peters, K. J., 2013. Evaluation of breeding objectives for purebred and crossbred selection schemes for adoption in indigenous chicken breeding programmes. Br. Poult. Sci. 54, 62–75. |
| [60] | Okumu, O. N., Ngeranwa, J. J. N., Binepal, Y. S., Kahi, A. K., Bramwel, W. W., Ateya, L. O., Wekesa, F. C., 2016. Genetic diversity of indigenous chickens from selected areas in Kenya using microsatellite markers. J. Genet. Eng. Biotechnol. |
| [61] | Padhi, M. K., 2016. Importance of Indigenous Breeds of Chicken for Rural Economy and Their Improvements for Higher Production Performance. Scientifica (Cairo). |
| [62] | Rutten, M. J. M., Bijma, P., Woolliams, J. A., Van Arendonk, J. A. M., 2002. SelAction: Software to predict selection response and rate of inbreeding in livestock breeding programs. J. Hered. 93, 456–458. |
| [63] | Saxena, V. K., Kolluri, G., 2018. Selection Methods in Poultry Breeding: From Genetics to Genomics, in: Application of Genetics and Genomics in Poultry Science. InTech. |
| [64] | Schrooten, C., Bovenhuis, H., Van Arendonk, J. A. M., Bijma, P., 2005. Genetic progress in multistage dairy cattle breeding schemes using genetic markers. J. Dairy Sci. 88, 1569–1581. |
| [65] | Shapiro, B. I., Gebru, G., Desta, S., Nigussie, K., 2017. Rwanda Livestock Master Plan Developed by the International Livestock Research Institute (ILRI) Livestock Master Plan (LMP) team. Kigali_Rwanda. |
| [66] | Sharmaa, A., Lee, J. S., Dang, C. G., Sudrajad, P., Kim, H. C., Yeon, S. H., Kang, H. S., Lee, S. H., 2015. Stories and challenges of genome wide association studies in livestock - a review. Asian-Australasian J. Anim. Sci. |
| [67] | Sitzenstock, F., Ytournel, F., Sharifi, A. R., Cavero, D., Täubert, H., Preisinger, R., Simianer, H., 2013. Efficiency of genomic selection in an established commercial layer breeding program. Genet. Sel. Evol. 45. |
| [68] | Smith, C., 1983. Estimated costs of genetic conservation in farm animals, in: Animal Genetic Resources Conservation by Management, Data Banks and Training. Proceedings of the Joint FAO-UNEP Expert Panel Meeting,. pp. 21–30. |
| [69] | Soller, M., Weigend, S., Romanov, M. N., Dekkers, J. C. M., Lamont, S. J., 2006. Strategies to assess structural variation in the chicken genome and its associations with biodiversity and biological performance. Poult. Sci. 85, 2061–2078. |
| [70] | Takele taye Desta, Dana, N., Ayalew, W., Hanotte, O., 2011. Current state of knowledge on phenotypic characteristics of indigenous chickens in the tropics. Worlds. Poult. Sci. J. 67, 507–516. |
| [71] | Täubert, H., Rensing, S., Reinhardt, F., 2011. Comparing conventional and genomic breeding programs with ZPLAN+. Interbull Bull. 0. |
| [72] | Thomasen, J. R., Liu, H., Sørensen, A. C., 2020. Genotyping more cows increases genetic gain and reduces rate of true inbreeding in a dairy cattle breeding scheme using female reproductive technologies. J. Dairy Sci. 103, 597–606. |
| [73] | Venkatas, J., Adeleke, M. A., Peters, S. O., Adebambo, O. A., 2019. Phenotypic Differentiation of Purebred and Crossbred Indigenous Chicken Genotypes Using Multivariate Analysis. Int. J. Poult. Sci. 18, 301–308. |
| [74] | Venkatramaiah, A., Mohapatra, S. C., Sinha, R., Ayyagari, V., Choudhuri, D., 1986. Selection response for part period egg number and egg mass in chickens - a comparison. Theor. Appl. Genet. 72, 129–134. |
| [75] | Villanueva B., Wrayh N.R., Thompson R. Prediction of asymptotic rates of response from selection on multiple traits using univariate and multivariate best linear unbiased predictors. Anim. Prod. 1993; 57: 1–13. |
| [76] | Wang, M. S., Zhang, R. W., Su, L. Y., Li, Y., Peng, M. S., Liu, H. Q., Zeng, L., Irwin, D. M., Du, J. L., Yao, Y. G., Wu, D. D., Zhang, Y. P., 2016. Positive selection rather than relaxation of functional constraint drives the evolution of vision during chicken domestication. Cell Res. 26, 556–573. |
| [77] | Weigel, K. A., 2001. Controlling Inbreeding in Modern Breeding Programs. J. Dairy Sci. 84, E177–E184. |
| [78] | Wellmann, R., Bennewitz, J., 2019. Key genetic parameters for population management. Front. Genet. 10, 1–20. |
| [79] | Wolc, A., Kranis, A., Arango, J., Settar, P., Fulton, J. E., O’Sullivan, N. P., Avendano, A., Watson, K. A., Hickey, J. M., de los Campos, G., Fernando, R. L., Garrick, D. J., Dekkers, J. C. M., 2016. Implementation of genomic selection in the poultry industry. Anim. Front. 6, 23–31. |
| [80] | Wondmeneh, E. H, Van der Waaij, D. T., Okeyo, M., van Arendonk, J. A. M., 2014. A running breeding program for indigenous chickens in Ethiopia: evaluation of success, in: Proceedings, 10th World Congress of Genetics Applied to Livestock Production. pp. 10–12. |
| [81] | Woolliams, J. A., Bijma, P., 2000. Predicting rates of inbreeding in populations undergoing selection. Genetics 154, 1851–64. |
| [82] | Woolliams, J. A., Bijma, P., Villanueva, B., 1999. Expected genetic contributions and their impact on gene flow and genetic gain. Genetics 153, 1009–1020. |
APA Style
Habimana, R., Ngeno, K., Okeno, T. O. (2024). Response to Selection of Indigenous Chicken in Rwanda Using Within-Breed Selection Strategy. Animal and Veterinary Sciences, 12(3), 95-106. https://doi.org/10.11648/j.avs.20241203.13
ACS Style
Habimana, R.; Ngeno, K.; Okeno, T. O. Response to Selection of Indigenous Chicken in Rwanda Using Within-Breed Selection Strategy. Anim. Vet. Sci. 2024, 12(3), 95-106. doi: 10.11648/j.avs.20241203.13
AMA Style
Habimana R, Ngeno K, Okeno TO. Response to Selection of Indigenous Chicken in Rwanda Using Within-Breed Selection Strategy. Anim Vet Sci. 2024;12(3):95-106. doi: 10.11648/j.avs.20241203.13
@article{10.11648/j.avs.20241203.13,
author = {Richard Habimana and Kiplangat Ngeno and Tobias Otieno Okeno},
title = {Response to Selection of Indigenous Chicken in Rwanda Using Within-Breed Selection Strategy
},
journal = {Animal and Veterinary Sciences},
volume = {12},
number = {3},
pages = {95-106},
doi = {10.11648/j.avs.20241203.13},
url = {https://doi.org/10.11648/j.avs.20241203.13},
eprint = {https://article.sciencepublishinggroup.com/pdf/10.11648.j.avs.20241203.13},
abstract = {The study evaluated response to selection from within-breed selection strategy for conventional (CBS) and genomic (GBS) breeding schemes. These breeding schemes were evaluated in both high-health environments (nucleus) and smallholder farms (commercial). Breeding goal was to develop a dual-purpose IC for both eggs and meat through selective breeding. Breeding objectives were body weight (BW), egg weight (EW), egg number (EN) and resistance to Newcastle disease (AbR). A deterministic simulation was performed to evaluate rates of genetic gain and inbreeding. Base population in the nucleus was made up of 40 cockerels and 200 pullets. Selection pressure was 4% and 20% in the males and the females, respectively. The impact of nucleus size and selection pressure on rates of genetic gain and inbreeding of the breeding program was investigated through sensitivity analysis. SelAction software was used to predict rates of genetic gain and inbreeding. Results showed that using CBS in the nucleus, the breeding goal was 340.41$ and 1.13 times higher than that in the commercial flock. Inbreeding rate per generation of selected chicken in the nucleus was 1.45% and lower by 1.32 times compared to their offspring under smallholder farms. Genetic gains per generation in the nucleus for BW and EN traits were 141.10 g and 1.07 eggs and 1.12 and 1.10 times greater than those in smallholder farms, respectively. With GBS, the breeding goal was increased by 3.00 times whereas inbreeding rate was reduced by 3.15 times. Besides, using GBS, the selected birds in the nucleus were relatively similar to those in a commercial environment. Finally, the study revealed that the nucleus size and mating ratio influence the rates of genetic gain and inbreeding in both GBS and CBS. This study shows that IC in Rwanda have the potential to be improved through within-breed selection strategy using either CBS or GBS.
},
year = {2024}
}
TY - JOUR T1 - Response to Selection of Indigenous Chicken in Rwanda Using Within-Breed Selection Strategy AU - Richard Habimana AU - Kiplangat Ngeno AU - Tobias Otieno Okeno Y1 - 2024/06/25 PY - 2024 N1 - https://doi.org/10.11648/j.avs.20241203.13 DO - 10.11648/j.avs.20241203.13 T2 - Animal and Veterinary Sciences JF - Animal and Veterinary Sciences JO - Animal and Veterinary Sciences SP - 95 EP - 106 PB - Science Publishing Group SN - 2328-5850 UR - https://doi.org/10.11648/j.avs.20241203.13 AB - The study evaluated response to selection from within-breed selection strategy for conventional (CBS) and genomic (GBS) breeding schemes. These breeding schemes were evaluated in both high-health environments (nucleus) and smallholder farms (commercial). Breeding goal was to develop a dual-purpose IC for both eggs and meat through selective breeding. Breeding objectives were body weight (BW), egg weight (EW), egg number (EN) and resistance to Newcastle disease (AbR). A deterministic simulation was performed to evaluate rates of genetic gain and inbreeding. Base population in the nucleus was made up of 40 cockerels and 200 pullets. Selection pressure was 4% and 20% in the males and the females, respectively. The impact of nucleus size and selection pressure on rates of genetic gain and inbreeding of the breeding program was investigated through sensitivity analysis. SelAction software was used to predict rates of genetic gain and inbreeding. Results showed that using CBS in the nucleus, the breeding goal was 340.41$ and 1.13 times higher than that in the commercial flock. Inbreeding rate per generation of selected chicken in the nucleus was 1.45% and lower by 1.32 times compared to their offspring under smallholder farms. Genetic gains per generation in the nucleus for BW and EN traits were 141.10 g and 1.07 eggs and 1.12 and 1.10 times greater than those in smallholder farms, respectively. With GBS, the breeding goal was increased by 3.00 times whereas inbreeding rate was reduced by 3.15 times. Besides, using GBS, the selected birds in the nucleus were relatively similar to those in a commercial environment. Finally, the study revealed that the nucleus size and mating ratio influence the rates of genetic gain and inbreeding in both GBS and CBS. This study shows that IC in Rwanda have the potential to be improved through within-breed selection strategy using either CBS or GBS. VL - 12 IS - 3 ER -
Department of Animal Production, University of Rwanda, Nyagatare, Rwanda; Department of Animal Science, Egerton University, Egerton, Kenya
Department of Agriculture and Natural Resources, Moi University, Eldoret, Kenya
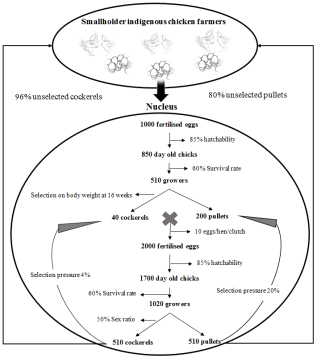
Figure 1. Breeding structure for the simulated indigenous chicken breeding program in Rwanda.
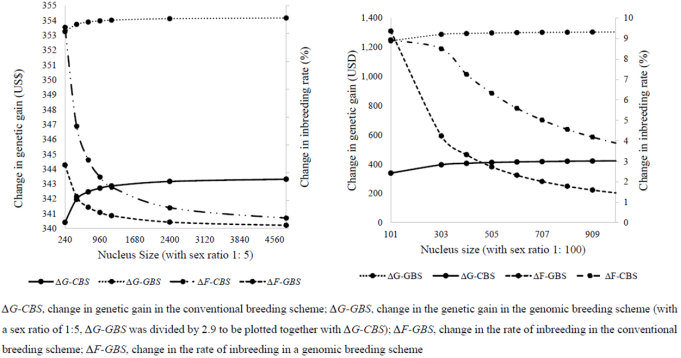
Figure 2. Trends for the rate of genetic gain and inbreeding attributed to the increase in nucleus size.
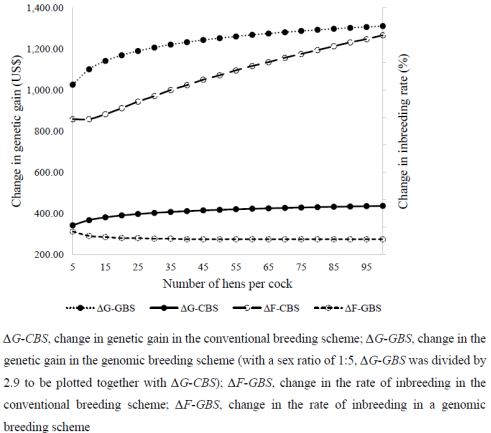
Figure 3. Trends for change in the rate of genetic gain and inbreeding due to the increase in the mating ratio.
Information

