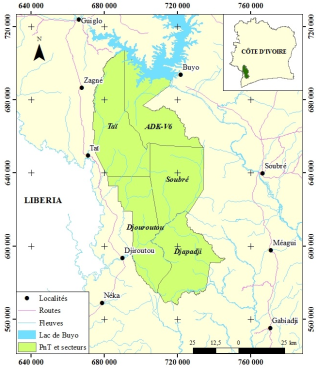
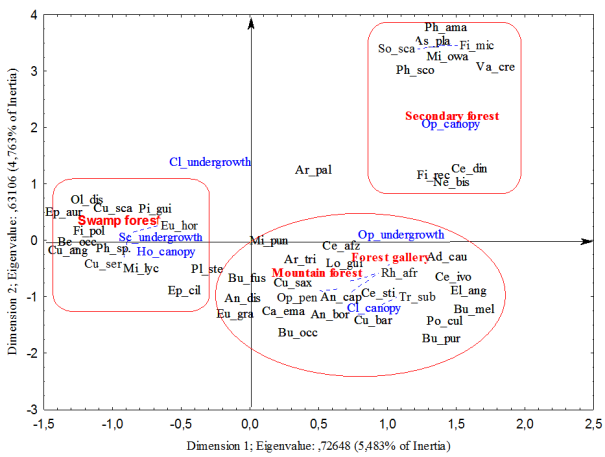
Anthropogenic pressures in the eastern zone of the Taï National Park have led to the fragmentation of plant formations. The forests in the eastern part of the park have suffered significant degradation of plant cover and a significant loss of biodiversity. Therefore, this study makes it possible to contribute to monitoring the dynamics of reconstitution of the plant cover based on the ecological characteristics and the distribution of epiphytes present in the Djapadji management sector. Floristic inventories were carried out. In the plots, all trees with a DBH ≥ 5 cm were counted and recorded for the study of the structure of plant formations, all species carrying an epiphyte were recorded and the epiphyte was identified. The analysis of the data presents a flora composed of 26 epiphytes distributed in 18 genera and 9 families, mainly present in mountain forests. The diversity of epiphytic plants is highest in mountain forests, followed by hydromorphic, secondary and gallery forests. Strict epiphytes and Hemiepiphytes are more present in the most preserved habitats, while accidental epiphytes are observed in reconstitution biotopes. The distribution of epiphytic plants allows us to affirm that the formerly anthropized forests of the Djapadji sector present a good dynamic of reconstitution.
| Published in | American Journal of Life Sciences (Volume 12, Issue 4) |
| DOI | 10.11648/j.ajls.20241204.11 |
| Page(s) | 65-72 |
| Creative Commons |
This is an Open Access article, distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution and reproduction in any medium or format, provided the original work is properly cited. |
| Copyright |
Copyright © The Author(s), 2024. Published by Science Publishing Group |
Epiphyte, Microhabitats, Biodiversity Conservation, Ecological Monitoring
Biotopes | Epiphytic Taxa | ||
|---|---|---|---|
Species | Genera | Families | |
Mountain Forests | 16 | 14 | 8 |
Secondary Forests | 9 | 8 | 8 |
Hydromorphic Forests | 10 | 9 | 7 |
Gallery forests | 14 | 11 | 5 |
№ | Taxon | Family | Epiphyte Type | Biological Type |
|---|---|---|---|---|
1 | Adientum sp | Pteridaceae | Accidental | H |
2 | Ancistrorhynchus capitatus (Lindl.) Summerh. | Orchidaceae | Strict | Ch |
3 | Angraecum distichum Lindl. | Orchidaceae | Strict | Ch |
4 | Arthropteris palisotii (Desv.) Alston | Oleandraceae | Hemiepiphyte | rh |
5 | Bulbophyllum fuscum Lindl. | Orchidaceae | Strict | Ch |
6 | Bulbophyllum purpurearhachys | Orchidaceae | Strict | Ch |
7 | Calyptrochilum emarginatum (Afzel. ex Sw.) Schltr. | Orchidaceae | Strict | Ch |
8 | Cercestis afzelii Schott | Araceae | Hemiepiphyte | Lmp |
9 | Cercestis dinklagei Engl. | Araceae | Hemiepiphyte | Lmp |
10 | Cercestis ivorensis A.Chev. | Araceae | Hemiepiphyte | Lmp |
11 | Cercestis stigmaticus N.E.Br. | Araceae | Hemiépiphyte | Lmp |
12 | Culcasia barombensis N.E.Br. | Araceae | Hemiepiphyte | Lmp |
13 | Culcasia saxatilis A.Chev. | Araceae | Hemiepiphyte | Lmp |
14 | Culcasia scandens P.Beauv. | Araceae | Hemiepiphyte | Lmp |
15 | Culcasia seretii De Wild. | Araceae | Hemiepiphyte | Lmp |
16 | Elaphoglossum angulatum (Blume) T.Moore | Dryopteridaceae | Strict | H |
17 | Eulophia gracilis Lindl. | Orchidaceae | Accidental | Ch |
18 | Eulophia horsfallii (Bateman) Summerh. | Orchidaceae | Accidental | Ch |
19 | Lomariopsis guineensis (Underw.) Alston | Lomariopsidaceae | Hemiepiphyte | G |
20 | Microsorum punctatum (L.) Copel. | Polypodiaceae | Strict | H |
21 | Nephrolepis biserrata (Sw.) Desv. | Nephrolepidaceae | Strict / Accidental | H |
22 | Philodendron sp | Araceae | Hemiepiphyte | Lmp |
23 | Piper guineense Schumach. & Thonn. | Piperaceae | Hemiepiphyte | Mp |
24 | Platycerium stemaria (P.Beauv.) Desv. | Polypodiaceae | Strict | H |
25 | Rhaphidophora africana N.E.Br. | Araceae | Strict | Lmp |
26 | Vanilla crenulata Rolfe | Orchidaceae | Hemiepiphyte | Lmp |
Biotopes | Shannon index | Pielou's index |
|---|---|---|
Mountain Forests | 2,53±0,09a | 0,89±0,42b |
Secondary Forests | 2,38±0,07a | 0,78±0,04a |
Hydromorphic Forests | 2,4±0,10a | 0,74±0,64a |
Gallery forests | 2,29±0,10a | 0,81±0,49b |
Mountain forests | Secondary forests | Hydromorphic forests | Forest galleries | |
|---|---|---|---|---|
Mountain forests | 100 | |||
Secondary forests | 40 | 100 | ||
Hydromorphic forests | 44.4 | 44.4 | 100 | |
Forest galleries | 56.3 | 52.2 | 56 | 100 |
APG | Angiosperm Phylogeny Group |
OIPR | Ivorian Office of Parks and Reserves |
CA | Correspondence Analysis |
MCA | Multiple Correspondence Analysis |
| [1] | Benzing, D. H., Vascular epiphytes. Forest canopies, 2004. 2: p. 175-211. |
| [2] | Zotz, G. and P. Hietz, The physiological ecology of vascular epiphytes: current knowledge, open questions. Journal of Experimental Botany, 2001. 52(364): p. 2067 - 2078. |
| [3] | Kelly, D. L., et al., The epiphyte communities of a montane rain forest in the Andes of Venezuela: patterns in the distribution of the flora. Journal of Tropical Ecology, 2004. 20(6): p. 643-666. |
| [4] | Zotz, G., et al., EpiList 1.0: a global checklist of vascular epiphytes. Ecology, 2021. 102(6): p. e03326. |
| [5] | Ibisch, P. I., et al., How diverse are neotropical epiphytes? An analysis based on the «Catalogue of the Flowering Plants and Gymnosperms of Peru». Ecotropica, 1996. 2: p. 13-28. |
| [6] | Nadkarni, N. M., Epiphyte biomass and nutrient capital of a neotropical elfin forest. Biotropica, 1984. 16(4): p. 249-256. |
| [7] | Gnagbo, A., Diversity, distribution and uses of vascular epiphytes from the lower strata of the coastal forests of Côte d’Ivoire: Case of Azagny National Park. 2015, Thesis, Félix HOUPHOUËT-BOIGNY University, Abidjan, Côte d'Ivoire. 165p. |
| [8] | Nadkarni, N. M., The conservation of epiphytes and their habitats: summary of a discussion at the international symposium on the biology and conservation of epiphytes. Selbyana, 1992. 13: p. 140-142. |
| [9] | Richter, M., Methoden der Klimmaindikation durch pflanzenmorphologische Merkmale in den Kordilleren der Neotropis. Die Erde, 1991. 122(4): p. 167-189. |
| [10] | Tsai, Y.-C., et al., Distinct epiphyte responses to drought in tropical mountain cloud forests. Ecohydrology, 2023. n/a(n/a): p. e2569. |
| [11] | Liu, Q., et al., Orchid diversity and distribution pattern in karst forests in eastern Yunnan Province, China. Forest Ecosystems, 2023. 10: p. 100117. |
| [12] | Dias-Pereira, J., et al., Vascular Epiphyte Diversity in a Key Atlantic Forest Remnant from Minas Gerais State, Southeastern Brazil. Floresta e ambiente, 2023. 30(1). |
| [13] | Taylor, A., et al., Vascular epiphytes contribute disproportionately to global centres of plant diversity. Global Ecology and Biogeography, 2022. 31(1): p. 62-74. |
| [14] | KÖSter, N., et al., Conservation of Epiphyte Diversity in an Andean Landscape Transformed by Human Land Use. Conservation Biology, 2009. 23(4): p. 911-919. |
| [15] | Kouamé, N. F., Influence of logging on the vegetation and flora of the Haut Sassandra classified forest (Central-West Côte d'Ivoire). 1998, Thesis, Cocody University, Abidjan, Côte d’Ivoire, 227 p. |
| [16] | Gnagbo, A., Phytogeography and uses of vascular epiphytes in Côte d’Ivoire”. 2019: European university publishing. 189 Pages. |
| [17] | OIPR, The universe of Parks and Reserves. OIPR Newsletter, n°1, 2007: p. 24 p. |
| [18] | Diarrassouba, A., et al., Differential response of seven duiker species to human activities in Taï National Park, Côte d'Ivoire. African Journal of Ecology, 2019. 58(1): p. 58-68. |
| [19] | Mangenot, G., Study on the forests of the plains and plateaus of Côte d'Ivoire. Eburnean studies. IFAN Dakar, 1956. tome 4: p. 55-67. |
| [20] | Bouko, B. S., B. Sinsin, and B. G. Soulé, Effects of land use dynamics on the structure and floristic diversity of open forests and savannahs in Benin. Tropicultura, 2007. 25(4): p. 221-227. |
| [21] | Aké-Assi, L., Flora of Côte d’Ivoire 1, systematic catalog, bio-geography and ecology. Conservatory and Botanical Garden, Geneva, Switzerland, 2001: 396 p. |
| [22] | Aké-Assi, L., Flora of Côte d’Ivoire 1, systematic catalog, bio-geography and ecology. Conservatory and Botanical Garden, Geneva, Switzerland, 2002: p. 401p. |
| [23] | Shannon, C. E., A Mathematical Theory of Communication. The Bell System Technical Journal 1948. 27: p. 379-423. |
| [24] | Felfili, J. M., et al., Diversity, floristic and structural patterns of cerrado vegetation in Central Brazil. Plant Ecology, 2004. 175(1): p. 37-46. |
| [25] | Hakizimana, P., Analysis of the composition, spatial structure and natural plant resources collected from the dense forest of Kigwena and the clear forest of Rumonge in Burundi. 2012, Doctoral thesis, Free University of Brussels, Belgium. p. 141. |
| [26] | Pielou, E. C., The measurement of diversity in different types of biological collections. Journal of theoretical biology, 1966. 13: p. 131-144. |
| [27] | Frontier, S., et al., Ecosystems. Structure, operation, evolution. 2008: Dunod, 4th edition, Paris. |
| [28] | Sorensen, T., A method of establishing groups of equal amplitude in plant sociology based on similarity of species content and its application to analysis of the vegetation on Danish commons. Biol. Skr., 1948. 5: p. 1-34. |
| [29] | Géhu, J.-M. and J. Géhu, Objection test for the biological assessment of natural environments. Coastal examples. Applied Phytosociology Seminar. French-speaking Phytosociology Association, Metz, 1980: p. 75-94. |
| [30] | Dajoz, R., Summary of ecology. 8th edition, Dunod, Paris, 2006: 631 p. |
| [31] | Palm, R., Multiple correspondence analysis: principles and application. Statistics and Computer Science Notes, 2007(2). |
| [32] | Moreau, J., P.-A. Doudin, and P. Cazes, Correspondence analysis and related techniques: new approaches for statistical data analysis. Vol. 32. 1999: Springer Science & Business Media. |
| [33] | Duval, J., Correspondence analysis and field construction. Proceedings of social science research, 2013(5): p. 110-123. |
| [34] | Gnagbo, A., D. Kouame, and C. Y. Adou Yao, Diversity of vascular epiphytes in the lower stratum of plant formations in Azagny National Park (South of Côte d’Ivoire). Journal of Animal &Plant Sciences, 2016. 28(1): p. 4366-4386. |
| [35] | Gnagbo, A., K. B. Kpangui, and C. Y. Adou Yao, Distribution of epiphytes in Côte d’Ivoire: effects of phytogeographic zones and rainfall variations. Afrique SCIENCE 2015. 11(1): p. 175-186. |
| [36] | Hubbell, S. P., The unified neutral theory of biodiversity and biogeography. Monographs in Population Biology, Princeton University Press, Princeton, 2001. Volume 32: 375 p. |
| [37] | Noumi, V. N., et al., Eco-biogeography of the Peperomia genus in Africa: case of the Guineo‐Congolese region. International Journal of Environmental Studies, 2011. 68(1): p. 43-60. |
APA Style
Anthelme, G., Mathieu, E. W., Junior, P. F. P., Lydie, K. M., Carmel, Y. K. K. A., et al. (2024). Epiphytic Dynamics of the Adaptations to the Changes of Habitats in Taï National Park. American Journal of Life Sciences, 12(4), 65-72. https://doi.org/10.11648/j.ajls.20241204.11
ACS Style
Anthelme, G.; Mathieu, E. W.; Junior, P. F. P.; Lydie, K. M.; Carmel, Y. K. K. A., et al. Epiphytic Dynamics of the Adaptations to the Changes of Habitats in Taï National Park. Am. J. Life Sci. 2024, 12(4), 65-72. doi: 10.11648/j.ajls.20241204.11
AMA Style
Anthelme G, Mathieu EW, Junior PFP, Lydie KM, Carmel YKKA, et al. Epiphytic Dynamics of the Adaptations to the Changes of Habitats in Taï National Park. Am J Life Sci. 2024;12(4):65-72. doi: 10.11648/j.ajls.20241204.11
@article{10.11648/j.ajls.20241204.11,
author = {Gnagbo Anthelme and Egnankou Wadja Mathieu and Pagny Frank Placide Junior and Kouao Marthe Lydie and Yao Koffi Kan Anicet Carmel and Tiébré Marie-Solange and Kouassi Kouadio Henri and Adou Yao Constant Yves},
title = {Epiphytic Dynamics of the Adaptations to the Changes of Habitats in Taï National Park
},
journal = {American Journal of Life Sciences},
volume = {12},
number = {4},
pages = {65-72},
doi = {10.11648/j.ajls.20241204.11},
url = {https://doi.org/10.11648/j.ajls.20241204.11},
eprint = {https://article.sciencepublishinggroup.com/pdf/10.11648.j.ajls.20241204.11},
abstract = {Anthropogenic pressures in the eastern zone of the Taï National Park have led to the fragmentation of plant formations. The forests in the eastern part of the park have suffered significant degradation of plant cover and a significant loss of biodiversity. Therefore, this study makes it possible to contribute to monitoring the dynamics of reconstitution of the plant cover based on the ecological characteristics and the distribution of epiphytes present in the Djapadji management sector. Floristic inventories were carried out. In the plots, all trees with a DBH ≥ 5 cm were counted and recorded for the study of the structure of plant formations, all species carrying an epiphyte were recorded and the epiphyte was identified. The analysis of the data presents a flora composed of 26 epiphytes distributed in 18 genera and 9 families, mainly present in mountain forests. The diversity of epiphytic plants is highest in mountain forests, followed by hydromorphic, secondary and gallery forests. Strict epiphytes and Hemiepiphytes are more present in the most preserved habitats, while accidental epiphytes are observed in reconstitution biotopes. The distribution of epiphytic plants allows us to affirm that the formerly anthropized forests of the Djapadji sector present a good dynamic of reconstitution.
},
year = {2024}
}
TY - JOUR T1 - Epiphytic Dynamics of the Adaptations to the Changes of Habitats in Taï National Park AU - Gnagbo Anthelme AU - Egnankou Wadja Mathieu AU - Pagny Frank Placide Junior AU - Kouao Marthe Lydie AU - Yao Koffi Kan Anicet Carmel AU - Tiébré Marie-Solange AU - Kouassi Kouadio Henri AU - Adou Yao Constant Yves Y1 - 2024/07/15 PY - 2024 N1 - https://doi.org/10.11648/j.ajls.20241204.11 DO - 10.11648/j.ajls.20241204.11 T2 - American Journal of Life Sciences JF - American Journal of Life Sciences JO - American Journal of Life Sciences SP - 65 EP - 72 PB - Science Publishing Group SN - 2328-5737 UR - https://doi.org/10.11648/j.ajls.20241204.11 AB - Anthropogenic pressures in the eastern zone of the Taï National Park have led to the fragmentation of plant formations. The forests in the eastern part of the park have suffered significant degradation of plant cover and a significant loss of biodiversity. Therefore, this study makes it possible to contribute to monitoring the dynamics of reconstitution of the plant cover based on the ecological characteristics and the distribution of epiphytes present in the Djapadji management sector. Floristic inventories were carried out. In the plots, all trees with a DBH ≥ 5 cm were counted and recorded for the study of the structure of plant formations, all species carrying an epiphyte were recorded and the epiphyte was identified. The analysis of the data presents a flora composed of 26 epiphytes distributed in 18 genera and 9 families, mainly present in mountain forests. The diversity of epiphytic plants is highest in mountain forests, followed by hydromorphic, secondary and gallery forests. Strict epiphytes and Hemiepiphytes are more present in the most preserved habitats, while accidental epiphytes are observed in reconstitution biotopes. The distribution of epiphytic plants allows us to affirm that the formerly anthropized forests of the Djapadji sector present a good dynamic of reconstitution. VL - 12 IS - 4 ER -
Department of Agroforestry, Jean Lorougnon Guédé University, Daloa, Côte d’Ivoire; Conservation and Valorization of Natural Resources, Swiss Center for Scientific Research, Abidjan, Côte d’Ivoire
Department of Biosciences, Félix Houphouët-Boigny University, Abidjan, Côte d’Ivoire
Department of Environment, Jean Lorougnon Guédé University, Daloa, Côte d’Ivoire
Conservation and Valorization of Natural Resources, Swiss Center for Scientific Research, Abidjan, Côte d’Ivoire
Department of Agroforestry, Jean Lorougnon Guédé University, Daloa, Côte d’Ivoire
Department of Biosciences, Félix Houphouët-Boigny University, Abidjan, Côte d’Ivoire; National Floristic Center, Félix Houphouët-Boigny University, Abidjan, Côte d’Ivoire
Department of Agroforestry, Jean Lorougnon Guédé University, Daloa, Côte d’Ivoire
Department of Biosciences, Félix Houphouët-Boigny University, Abidjan, Côte d’Ivoire
Information