The mobilization of teneral reserve components across mosquito life stages (eggs-pupae) is vital because these teneral components are required for disease transmission. This study was conducted to determine the influence of teneral reserve accumulation on wing development and vectoral fitness in Anopheles gambiae and Culex quinquefasciatus mosquitoes. The first live instar (L1) of the selected mosquitoes was sampled using a 350 ml dipper from larval habitats that included gutters, swamps, and large water bodies of the selected sites in Minna metropolis, reared to the fourth live instar (L4), pupae, and adults under laboratory conditions of temperature and relative humidity (28°C and 73%, respectively) in separate modified larval and dried adult holding cages (LHC and DAHC). At the adult stage, the wings of mosquitoes were removed and measured under a microscope, and nutritional reserves and vectorial fitness were determined using measured wings as an index. Differences in the nutritional composition of the fourth instar An. gambiae were as follows: sugar (3.66±0.20 microgram/larvae), glycogen (2.09±0.26 µg/lar), lipid (8.50±1.0 µg/lar) and proteins (22.39±0.63 µg/lar). Cx. quinquefasciatus had teneral reserves as follows (4.14±0.30, 2.07±0.28, 10.00±0.68 and 22.56±0.46 µg/Lar) of sugar, glycogen, lipid, and protein respectively. The dissimilarity in wing length among species populations emerged only within a narrow range. The relatively larger wings were visible in the average wing length of Cx quinquefasciatus (3.77±0.12 millimetre). The proportion of wings of both left and right wings were 0.03±0.01 mm among the two different species populations. In addition, significant positive to highly positive correlations were observed between nutritional components and wing length, sugar (r= 0.489), protein (r=0.991**), and wing length in An. gambiae, whereas a negative correlation was observed for all teneral content in Cx. quiquefasciatus (r= -0.112 to -0.830). These findings were evaluated for the suitability of disease spread and are expected to contribute to the development of anti-larval control strategies through the manipulation of larval habitat nutrient contents.
| Published in | American Journal of Entomology (Volume 9, Issue 2) |
| DOI | 10.11648/j.aje.20250902.14 |
| Page(s) | 87-99 |
| Creative Commons |
This is an Open Access article, distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution and reproduction in any medium or format, provided the original work is properly cited. |
| Copyright |
Copyright © The Author(s), 2025. Published by Science Publishing Group |
Teneral Reserves, Larval Instar, Wing Length, Vectoral Fitness, Nutritional Composition
Food Nutrients | ||||
|---|---|---|---|---|
Sites | Sugar (µg/Lar) | Glycogen (µg/Lar) | Lipid (µg/Lar) | Protein (µg/Lar) |
A | 3.60±0.23ab | 3.66±0.22c | 14.66±1.33c | 24.49±0.93b |
B | 4.06±0.35b | 1.21±0.11a | 8.67±0.82a | 24.03±0.61b |
C | 4.14±0.50b | 2.15±0.54b | 5.33±1.33a | 19.46±0.60a |
D | 2.82±0.17a | 1.35±0.14a | 5.33±0.82a | 21.56±1.40a |
Mean | 3.66±0.20 | 2.09±0.26 | 8.50±1.01 | 22.39±0.63 |
Food Nutrients | ||||
|---|---|---|---|---|
Sites | Sugar (µg/Lar) | Glycogen (µg/Lar) | Lipid (µg/Lar) | Protein (µg/Lar) |
A | 5.70±0.60c | 3.97±0.34b | 11.34±2.26a | 24.34±0.68b |
B | 4.60±0.36b | 1.56±0.24a | 10.00±1.05a | 23.13±0.83ab |
C | 3.38±0.16a | 1.63±0.19a | 8.67±0.82a | 21.68±0.67a |
D | 2.88±0.13a | 1.10±0.04a | 10.00±1.05a | 21.11±0.92a |
Mean | 4.14±0.30 | 2.07±0.28 | 10.00±0.68 | 22.56±0.46 |
RWL (mm) | LWL (mm) | MWL (mm) | FA (mm) | |
|---|---|---|---|---|
Sugar | 0.407 | 0.423 | 0.489 | 0.884 |
Glycogen | -0.071 | -0.119 | -0.070 | -0.242 |
Lipid | -0.230 | -0.200 | -0.136 | 0.746 |
Protein | 0.974* | 0.978* | 0.991** | 0.600 |
RWL (mm) | LWL (mm) | MWL (mm) | FA (mm) | |
|---|---|---|---|---|
Sugar | -0.804 | -0.797 | -0.830 | 0.959* |
Glycogen | -0.648 | -0.650 | -0.658 | 0.012 |
Lipid | -0.746 | -0.753 | -0.701 | 0.228 |
Protein | -0.103 | -0.107 | -0.112 | -0.525 |
FCTA | Federal Capital Territory Abuja |
LGA | Local Government Area |
An. | Anopheline |
Cx | Culex |
L1 | Larval Stage 1 |
L2 | Larval Stage 2 |
L3 | Larval Stage 3 |
L4 | Larval Stage 4 |
LHC | Larval Holding Cages |
DAHC | Dry Adult Holding Cages |
OD | Optical Density |
RWL1 | Right Wing Length |
LWL1 | Left Wing Length |
MWL | Wing Length |
FA1 | Fluctuating Asymmetry |
| [1] | Kang, D. S., Alcalay, Y., Lovin, D. D., Cunningham, J. M., Eng, M. W., Chadee, D. D., & Severson, D. W. (2017). Larval stress alters dengue virus susceptibility in adult female Aedes aegypti (L.) adult females. Acta tropica, (174): 97-101. |
| [2] | Sasmita, H. I., Tu, W.-C., Bong, L.-J., & Neoh, K. B. (2019). Effects of larval diets and temperature regimes on life history traits, energy reserves and temperature tolerance of male Aedes aegypti (Diptera: Culicidae): optimizing rearing techniques for the sterile insect programmes. Parasites & vectors, 12(1): 1-16. |
| [3] | Mohan, S., Banerjee, S., Pramanik, S., Banerjee, S., Saha, G. K., & Aditya, G. (2017). Comparative account of energy reserves in four co-occurring mosquito species in Kolkata, India (Diptera: Culicidae). Polish Journal of Entomology, 86(1): 49. |
| [4] | Sultana, N., Khan, M., Karim, M. R., & Khatun, N. (2021). Efficiency of two larval diets for mass rearing of Aedes aegypti in support of sterile insect technique (SIT). Vector Research and Parasitology ISSN 2054-9881: Vol (8)1 |
| [5] | Ukubuiwe, A. C., Olayemi, I. K., Omalu, I. C. J., Arimoro, F. O., Baba, B. M., & Ukubuiwe, C. C. (2018). Influence of variable photoperiod on life-stages mobilization of teneral reserves in Culex quinquefasciatus (Diptera: Culicidae): implication for environmental manipulation for vector control. Molecular Entomology (9) 1: 1-10 |
| [6] | Zirbel, K. E., & Alto, B. W. (2018). Maternal and paternal nutrition in a mosquito influences offspring life histories but not infection with an arbovirus. Ecosphere, (9), 10. |
| [7] | Nascimento, D. A. S., Trindade, F. T. T., & Silva, A. d. A. e. (2021). Dietary supplementation with vitamins and minerals improves larvae and adult rearing conditions of Anopheles darlingi (Diptera: Culicidae). Journal of Medical Entomology, 58(1): 71-78. |
| [8] | Yan, J., Kibech, R., & Stone, C. M. (2021). Differential effects of larval and adult nutrition on female survival, fecundity, and size of the yellow fever mosquito, Aedes aegypti. Frontiers in Zoology, 18(1): 1-9. |
| [9] | Oyibo-Usman, K., Olayemi, I., Adebola, M., & Salaudeen, M. (2019). Evaluation of stem borers teneral reserve concentration and their effects on proximate composition and yield parameter of maize in agro-ecological Zone b, Niger State, Nigeria. International Journal of Applied Biological Research, 10(1): 145- 158. |
| [10] | Clifton, M. E., Correa, S., Rivera-Perez, C., Nouzova, M., & Noriega, F. G. (2014). Male Aedes aegypti mosquitoes use JH III transferred during copulation to influence previtellogenic ovary physiology and affect the reproductive output of female mosquitoes. Journal of Insect Physiology, (64): 40-47. |
| [11] | Barreaux, A. M., Stone, C. M., Barreaux, P., & Koella, J. C. (2018). The relationship between size and longevity of the malaria vector Anopheles gambiae (ss) depends on the larval environment. Parasites & Vectors, 11(1): 1-9. |
| [12] | soe, J., Koch, L. E., Isoe, Y. E., Rascón Jr, A. A., Brown, H. E., Massani, B. B., & Miesfeld, R. L. (2019). Identification and characterization of a mosquito-specific eggshell organizing factor in Aedes aegypti mosquitoes. PLoS Biology, 17(1). |
| [13] | Khan, I., Farid, A., and Zeb, A. (2013). Development of inexpensive and globally available larval diet for rearing Anopheles stephensi (Diptera: Culicidae) mosquitoes. Parasites & Vectors, 6(1) 1-7. |
| [14] | Briegel H. (1990). Metabolic relationship between female body size, reserves, and fecundity of Aedes aegypti, Journal of Insect Physiology, 36: 165–172 |
| [15] | Brault, A. C. (2009). Changing patterns of West Nile virus transmission: altered vector competence and host susceptibility. Vector Resistance (40)43 |
| [16] | Ibrahim, S. K. (2022). Species composition and productivity of mosquitoes in relation to physico-chemical conditions, general reserves, insecticide susceptibility and community knowledge on mosquito-borne diseases in Niger, Nigeria. PhD thesis, Approved by the Institute of Postgraduate School, Universiti Sains Malaysia. Pg 47-51. |
| [17] | Shehu, I. K., Bawa, S. M., Mohammed, A., Yakudima, I. I. & Hasber, S. (2023). Relative productivity of mosquito larval habitats for abundance and immature stages distribution in relation to physicochemical properties in Niger State, Nigeria. Nigerian Journal of Parasitology 44(1): 155-168 |
| [18] | Akpan, G., Adepoju, K., Oladosu, O., & Adelabu, S. (2018). Dominant malaria vector species in Nigeria: Modelling potential distribution of Anopheles gambiae sensu lato and its siblings with MaxEnt. PloS One, 13(10). |
| [19] | I. Kura Shehu, H. B., Ahmad, I. Kayode Olayemi, S. Danjuma, H. Abu Ahmad and & Hasber (2023). Insecticide susceptibility status in two medically important mosquito vectors; Anopheles gambiae and Culex quinquefasciatus to three insecticides commonly used in Niger State, Nigeria. Saudi Journal of Biological Sciences 30 (2023): 103524 |
| [20] | Wang, J.-N., Hou, J., Zhong, J.-Y., Cao, G.-P., Yu, Z.-Y., Wu, Y.-Y.,... and Gong, Z.-Y. (2020). Relationships between traditional larval indices and meteorological factors with the adult density of Aedes albopictus captured by BG-mosquito trap. PloS One, 15(6). |
| [21] | Ibrahim, K. S., Hamdan, A., Intan. H. I., Israel, K. O., Danjuma, S., Abu. H. A., Yakudima, I. I., Rashidu, M., & Hasber, S. (2022). Monthly Assessment of the relative abundance and development of immature stages of Culex mosquitoes (Diptera: Culicidae) of the selected breeding habitats in Niger, Nigeria. Serangga, 27(3): 187-203 ISSN 1394-5130 |
| [22] | Shehu, I. K., Ahmad, H., Olayemi, I. K., Solomon, D., Ahmad, A. H. & Salim, H. (2022). Molecular characterization of spatially heterogeneous populations of the malaria vector (Anopheles gambiae s.l) in Niger State, Nigeria. Nigerian Journal of Parasitology, 43(1) 47-57 |
| [23] | Babale, S. K., Salim, H., Yakudima, I. I., Kabir, B. M., Mamman, R., Chiroma, U. M., & Ibrahim, S. K. (2023). Seasonal assessment of pupal habitat productivity of malaria vector: Anopheles gambiae s.l as influence by physico-chemical conditions at selected breeding habitats in Niger, Nigeria. EUREKA: Life Sciences, 2, 38–55. |
| [24] | Kaufmann, C., & Brown, M. R. (2008). Regulation of carbohydrate metabolism and flight performance by a hypertrehalosaemic hormone in the mosquito Anopheles gambiae. Journal of Insect Physiology, 54(2), 367-377. |
| [25] |
Bova, J., Paulson, S., & Paulson, G. (2016). Morphological differentiation of the eggs of North American container-inhabiting Aedes mosquitoes. Journal of the American Mosquito Control Association, 32(3), 244-247. Downloaded From:
https://bioone.org/journals/Journal-of-the-American-Mosquito-Control-Association on 1/30/2019 |
| [26] | Niang, E. H. A., Bassene, H., Fenollar, F., & Mediannikov, O. (2018). Biological control of mosquito-borne diseases: the potential of Wolbachia-based interventions in an (IVM) framework. Journal of Tropical Medicine, 1470459. |
| [27] | Day, J. F. (2016). Mosquito oviposition behaviour and vector control. Insects, 7(4), 65. |
| [28] | Ukubuiwe, A., Olayemi, I., Omalu, I., Jibrin, A., & Oyibo-Usman, K. (2013). Molecular bases of reproductive and vectorial fitness of Culex pipiens pipiens (Diptera: Culicidae) mosquito populations, for the transmission of filariasis in North Central Nigeria. Journal of Medical Sciences, 13(3), 201-207. |
| [29] | Huck, D. T., Klein, M. S., & Meuti, M. E. (2021). Determining the effects of nutrition on the reproductive physiology of male mosquitoes. Journal of Insect Physiology, (129), 104191. |
| [30] |
D'Souza, J., & Nderitu, D. (2021). Ethical considerations for introducing RTS, S/AS01 in countries with moderate to high Plasmodium falciparum malaria transmission. The Lancet Global Health, 9(12). w
ww.thelancet.com/lancetgh Vol 9 December 2021. |
| [31] | Torre, A. (2015). Candidate biomarkers for mosquito age-grading identified by label-free quantitative analysis of protein expression in Aedes albopictus females. Journal of proteomics, (128), 272-279. |
| [32] |
Aminuwa, H., Olayemi, I. K., Ukubuiwe, A. C., Adeniyi, K. A., & Odeyemi, M. O. (2018). Evaluation of critical larval habitat physico-chemical factors on embryonic development and adult fitness of Culex quinquefasciatus mosquitoes (Diptera: Culicidae). Malaya Journal of Biosciences 2018, 5(2): 48-56.
www.malayabiosciences.com ISSN 2348-6236 print /2348-3075 online. |
APA Style
Kura, S. I., Salim, H., Hassan, A. A., Yakudima, I. I., Abdulazeez, A. K., et al. (2025). The Influence of Teneral Reserves Mobilization on Wing Development for Vectoral Fitness in Anopheles Gambiae and Culex Quinquefasciatus Mosquitoes. American Journal of Entomology, 9(2), 87-99. https://doi.org/10.11648/j.aje.20250902.14
ACS Style
Kura, S. I.; Salim, H.; Hassan, A. A.; Yakudima, I. I.; Abdulazeez, A. K., et al. The Influence of Teneral Reserves Mobilization on Wing Development for Vectoral Fitness in Anopheles Gambiae and Culex Quinquefasciatus Mosquitoes. Am. J. Entomol. 2025, 9(2), 87-99. doi: 10.11648/j.aje.20250902.14
AMA Style
Kura SI, Salim H, Hassan AA, Yakudima II, Abdulazeez AK, et al. The Influence of Teneral Reserves Mobilization on Wing Development for Vectoral Fitness in Anopheles Gambiae and Culex Quinquefasciatus Mosquitoes. Am J Entomol. 2025;9(2):87-99. doi: 10.11648/j.aje.20250902.14
@article{10.11648/j.aje.20250902.14,
author = {Shehu Ibrahim Kura and Hasber Salim and Ahmad Abu Hassan and Ismaila Ibrahim Yakudima and Adeniyi Kamoru Abdulazeez and Ibrahim Kabir Kontagora and Aminuwa Hyelamada Abuh and Shitta Kefas Babale and Saadatu Bawa and Buda Mohammed Kabir and Audu Dalladi Passi and Olayemi Isreal Kayode and Danjuma Solomon},
title = {The Influence of Teneral Reserves Mobilization on Wing Development for Vectoral Fitness in Anopheles Gambiae and Culex Quinquefasciatus Mosquitoes
},
journal = {American Journal of Entomology},
volume = {9},
number = {2},
pages = {87-99},
doi = {10.11648/j.aje.20250902.14},
url = {https://doi.org/10.11648/j.aje.20250902.14},
eprint = {https://article.sciencepublishinggroup.com/pdf/10.11648.j.aje.20250902.14},
abstract = {The mobilization of teneral reserve components across mosquito life stages (eggs-pupae) is vital because these teneral components are required for disease transmission. This study was conducted to determine the influence of teneral reserve accumulation on wing development and vectoral fitness in Anopheles gambiae and Culex quinquefasciatus mosquitoes. The first live instar (L1) of the selected mosquitoes was sampled using a 350 ml dipper from larval habitats that included gutters, swamps, and large water bodies of the selected sites in Minna metropolis, reared to the fourth live instar (L4), pupae, and adults under laboratory conditions of temperature and relative humidity (28°C and 73%, respectively) in separate modified larval and dried adult holding cages (LHC and DAHC). At the adult stage, the wings of mosquitoes were removed and measured under a microscope, and nutritional reserves and vectorial fitness were determined using measured wings as an index. Differences in the nutritional composition of the fourth instar An. gambiae were as follows: sugar (3.66±0.20 microgram/larvae), glycogen (2.09±0.26 µg/lar), lipid (8.50±1.0 µg/lar) and proteins (22.39±0.63 µg/lar). Cx. quinquefasciatus had teneral reserves as follows (4.14±0.30, 2.07±0.28, 10.00±0.68 and 22.56±0.46 µg/Lar) of sugar, glycogen, lipid, and protein respectively. The dissimilarity in wing length among species populations emerged only within a narrow range. The relatively larger wings were visible in the average wing length of Cx quinquefasciatus (3.77±0.12 millimetre). The proportion of wings of both left and right wings were 0.03±0.01 mm among the two different species populations. In addition, significant positive to highly positive correlations were observed between nutritional components and wing length, sugar (r= 0.489), protein (r=0.991**), and wing length in An. gambiae, whereas a negative correlation was observed for all teneral content in Cx. quiquefasciatus (r= -0.112 to -0.830). These findings were evaluated for the suitability of disease spread and are expected to contribute to the development of anti-larval control strategies through the manipulation of larval habitat nutrient contents.
},
year = {2025}
}
TY - JOUR T1 - The Influence of Teneral Reserves Mobilization on Wing Development for Vectoral Fitness in Anopheles Gambiae and Culex Quinquefasciatus Mosquitoes AU - Shehu Ibrahim Kura AU - Hasber Salim AU - Ahmad Abu Hassan AU - Ismaila Ibrahim Yakudima AU - Adeniyi Kamoru Abdulazeez AU - Ibrahim Kabir Kontagora AU - Aminuwa Hyelamada Abuh AU - Shitta Kefas Babale AU - Saadatu Bawa AU - Buda Mohammed Kabir AU - Audu Dalladi Passi AU - Olayemi Isreal Kayode AU - Danjuma Solomon Y1 - 2025/06/18 PY - 2025 N1 - https://doi.org/10.11648/j.aje.20250902.14 DO - 10.11648/j.aje.20250902.14 T2 - American Journal of Entomology JF - American Journal of Entomology JO - American Journal of Entomology SP - 87 EP - 99 PB - Science Publishing Group SN - 2640-0537 UR - https://doi.org/10.11648/j.aje.20250902.14 AB - The mobilization of teneral reserve components across mosquito life stages (eggs-pupae) is vital because these teneral components are required for disease transmission. This study was conducted to determine the influence of teneral reserve accumulation on wing development and vectoral fitness in Anopheles gambiae and Culex quinquefasciatus mosquitoes. The first live instar (L1) of the selected mosquitoes was sampled using a 350 ml dipper from larval habitats that included gutters, swamps, and large water bodies of the selected sites in Minna metropolis, reared to the fourth live instar (L4), pupae, and adults under laboratory conditions of temperature and relative humidity (28°C and 73%, respectively) in separate modified larval and dried adult holding cages (LHC and DAHC). At the adult stage, the wings of mosquitoes were removed and measured under a microscope, and nutritional reserves and vectorial fitness were determined using measured wings as an index. Differences in the nutritional composition of the fourth instar An. gambiae were as follows: sugar (3.66±0.20 microgram/larvae), glycogen (2.09±0.26 µg/lar), lipid (8.50±1.0 µg/lar) and proteins (22.39±0.63 µg/lar). Cx. quinquefasciatus had teneral reserves as follows (4.14±0.30, 2.07±0.28, 10.00±0.68 and 22.56±0.46 µg/Lar) of sugar, glycogen, lipid, and protein respectively. The dissimilarity in wing length among species populations emerged only within a narrow range. The relatively larger wings were visible in the average wing length of Cx quinquefasciatus (3.77±0.12 millimetre). The proportion of wings of both left and right wings were 0.03±0.01 mm among the two different species populations. In addition, significant positive to highly positive correlations were observed between nutritional components and wing length, sugar (r= 0.489), protein (r=0.991**), and wing length in An. gambiae, whereas a negative correlation was observed for all teneral content in Cx. quiquefasciatus (r= -0.112 to -0.830). These findings were evaluated for the suitability of disease spread and are expected to contribute to the development of anti-larval control strategies through the manipulation of larval habitat nutrient contents. VL - 9 IS - 2 ER -
Department of Science Education (Biology Unit), Abdulkadir Kure University, Minna, Nigeria
School of Biological Sciences, Universiti Sains Pulau, Penang, Malaysia
School of Biological Sciences, Universiti Sains Pulau, Penang, Malaysia
Department of Geography, Aliko Dangote University of Science and Technology, Wudil, Kano, Nigeria
Department of Animal and Environmental Biology, Federal University Dutse, Jigawa, Nigeria
Department of Agricultural Education, Federal University of Education, Kontagora, Nigeria
Department of Zoology, Ahmadu Bello University, Zaria, Nigeria
Department of Zoology, Federal University, Lokoja, Nigeria
Department of Biology, Niger State College of Education, Minna, Nigeria
Department of Biology, Niger State College of Education, Minna, Nigeria
Department of Biology, Federal University of Technology, Minna, Nigeria
Department of Crop Production, Ibrahim Badamasi Babangida University, Lapai, Nigeria
Figure 1. Map of Niger State showing the study area.
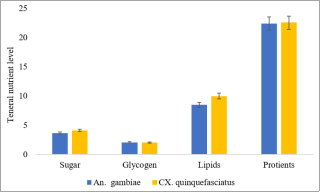
Figure 2. Mean distribution of teneral nutrients (µg/mosquito) on the fourth instar of An. gambaie and Cx. quinquefasciatus mosquito population.
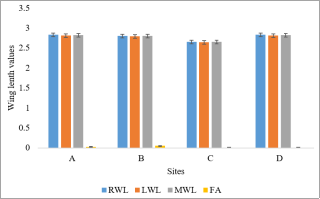
Figure 3. Wing lengths and fluctuating asymmetry of An. gambiae from different studies sites.
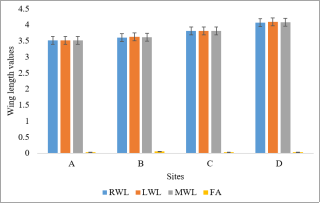
Figure 4. Wing lengths and fluctuating asymmetry of Cx. quinquefasciatus from selected studies sites.
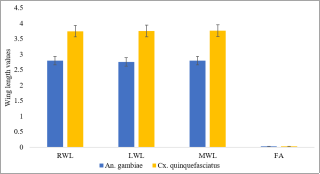
Figure 5. Mean wing length (mm) and asymmetry of adults An. gambiae and Cx. quinquefasciatus population.
Information

