Synthetic pesticides present worldwide risks of contamination of humans, livestock and the environment due to the strong persistence and the toxic residues in fruits and vegetables. Natural biopesticides of local plant origin present low persistence and are the best alternative for the control of crop pests. In the Adamaoua region (Northern Cameroon), few studies exist concerning effects of botanical pesticides on the behavior of beneficial insects. Studies aimed to draw up a list of pollinating insects on flowers of Vigna unguiculata (L.) Walp., 1843 (Fabales: Fabaceae), in situations of treatment with botanical pesticides compared to the situation of the use of synthetic insecticide and to determine the effect of the biopesticides on the behavior of the main floricultural insects. Field investigations were carried out during two cowpea cultivation campaigns (June to September 2021 and June to October 2022) in Dang (suburb of Ngaoundere) on the effect of leaves extracts of local plant origin on the foraging behavior of Apis mellifera Linnaeus, 1758 (Hymenoptera: Apidae) and the main sap-sucking insect Aphis craccivora Koch, 1854 (Hemiptera: Aphididae). Forty-four cowpea plots of 4x3.5 m each distributed according to the randomized complete block model (four untreated plots as negative control, four plots treated with the synthetic insecticide Parastar (40EC 535/ 10/IN, 20 g/l of imidacloprid and 20 g/l of lamda-cyhalothrin) as positive control, and 36 experimental plots treated with three concentrations (10%, 20% and 30%) of aqueous leaves extract of Calotropis procera (Gentianales: Apocynaceae), Eucalyptus camaldulensis (Myrtales: Myrtaceae), and Tithonia diversifolia (Asterales: Asteraceae) respectively, made it possible to conduct four treatments: (1) flowers left to freely pollination, (2) flowers protected against pollinators, (3) flowers visited exclusively by Ap. mellifera and (4) flowers protected against insects. Among eight species (four orders, four families and seven genera) recorded on the flowers of V. unguiculata, the domestic bee Ap. mellifera was the most common and collected nectar and pollen. The control plots and those treated with 10% or 20% aqueous leaves extracts allowed the bee to carry out its activity. Plots treated with 30% extract of each plant and those treated with the synthetic insecticide Parastar, drastically altered the rhythm and speed of activity in Ap. mellifera foragers. This behavior became less coordinated and slow on treated plants. It would be wise to use 10% or 20% aqueous extracts as botanical insecticides and an alternative to the synthetic insecticide Parastar.
| Published in | American Journal of Entomology (Volume 8, Issue 2) |
| DOI | 10.11648/j.aje.20240802.12 |
| Page(s) | 31-59 |
| Creative Commons |
This is an Open Access article, distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution and reproduction in any medium or format, provided the original work is properly cited. |
| Copyright |
Copyright © The Author(s), 2024. Published by Science Publishing Group |
Floricultural Insects, Vigna unguiculata, Botanical insecticides, Chemical Pest Control
Orders / Famillies / Species | Products | Collection periods | |||
|---|---|---|---|---|---|
2021: n (%) | 2022: n (%) | Total (%) | |||
Hemiptera Linnaeus, 1758 / Aphididae Latreille, 1802 | |||||
Aphis craccivora Koch, 1854 | Sap-sucking | 1,611 (17.9) | 3,003 (33.4) | 4,614 (51.3) | |
Hymenoptera Linnaeus, 1758 / Apidae Latreille, 1802 | |||||
Amegilla calens (Le Peletier, 1841) | Nectar | 199 (2.2) | 61 (0.7) | 260 (2.9) | |
Amegilla sp. Friese, 1897 | Nectar | 338 (3.8) | - | 338 (3.8) | |
Apis mellifera Linnaeus, 1758 | Nectar, and pollen | 1,338 (14.9) | 976 (10.9) | 2,314 (25.7) | |
Xylocopa olivacea (Fabricius 1778) | Nectar | 501 (5.6) | 236 (2.6) | 737 (8.2) | |
Lepidoptera Linnaeus, 1758 / Nymphalidae Rafinesque, 1815 | |||||
Danaus plexippus (Linnaeus, 1758) | Nectar | 94 (1.0) | - | 94 (1.0) | |
Hypolimnas misippus (Linnaeus, 1764) | Nectar | 69 (0.8) | 12 (0.1) | 81 (0.9) | |
Orthoptera Latreille, 1793 / Acridae Macleay, 1821 | |||||
Tettigonia viridissima (Linnaeus, 1758) | Phytophagous | 242 (2.7) | 307 (3.4) | 549 (6.1) | |
Total | 4,392 (48.9) | 4,595 (51.1) | 8,987 (100.0) | ||
A. Duration (s) of the nectar collection | B. Duration (s) of the pollen collection | A vs. B | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
n | Min. | Max. | Median | Mean ± se | n | Min. | Max. | Median | Mean ± se | Student t-test | |
2021 campaign | |||||||||||
Untreated | 145 | 1.0 | 15.0 | 10 | 9.3±0.3 | 140 | 2.0 | 10.0 | 7 | 6.4±0.2 | t=8.46, df=283, p<0.001* |
Parastar | 156 | 2.0 | 10.0 | 6 | 5.9±0.2 | 141 | 1.0 | 6.0 | 4 | 3.8±0.1 | t=10.01, df=295, p<0.001* |
Cp10 | 152 | 1.0 | 15.0 | 7 | 7.1±0.3 | 147 | 2.0 | 10.0 | 4 | 3.7±0.1 | t=11.53, df=297, p<0.001* |
Cp20 | 193 | 1.0 | 20.0 | 9 | 7.5±0.3 | 178 | 1.0 | 10.0 | 4 | 4.8±0.2 | t=7.05, df=369, p<0.001* |
Cp30 | 191 | 1.0 | 15.0 | 7 | 7.0±0.2 | 186 | 2.0 | 10.0 | 4 | 3.7±0.1 | t=12.87, df=375, p<0.001* |
Ec10 | 211 | 1.0 | 20.0 | 8 | 7.9±0.2 | 203 | 2.0 | 11.0 | 4 | 4.7±0.2 | t=11.37, df=412, p<0.001* |
Ec20 | 198 | 1.0 | 20.0 | 9 | 9.0±0.3 | 190 | 2.0 | 9.0 | 5 | 5.4±0.2 | t=10.43, df=386, p<0.001* |
Ec30 | 195 | 1.0 | 15.0 | 7 | 6.9±0.2 | 187 | 2.0 | 10.0 | 3 | 3.7±0.1 | t=12.85, df=380, p<0.001* |
Td10 | 138 | 1.0 | 15.0 | 7 | 7.0±0.3 | 133 | 2.0 | 10.0 | 4 | 3.7±0.1 | t=10.92, df=269, p<0.001* |
Td20 | 153 | 2.0 | 15.0 | 10 | 8.3±0.3 | 138 | 1.0 | 9.0 | 6 | 5.8±0.2 | t=6.98, df=289, p<0.001* |
Td30 | 103 | 1.0 | 15.0 | 7 | 6.6±0.3 | 98 | 2.0 | 10.0 | 4 | 3.7±0.2 | t=8.22, df=199, p<0.001* |
Pooled plots | 1835 | 1.0 | 20.0 | 8 | 7.5±3.5 | 1741 | 1.0 | 11.0 | 4 | 4.5±2.3 | t=30.7, df=3574, p<0.001* |
ANOVA | F(10; 1,824)=15.137, p<0.001 * | F(10; 1,824)=31.688, p<0.001 * | |||||||||
2022 campaign | |||||||||||
Untreated | 113 | 1.0 | 15.0 | 7 | 7.2±0.3 | 108 | 2.0 | 8.0 | 4 | 3.9±0.2 | t=10.40, df=219, p<0.001* |
Parastar | 128 | 2.0 | 10.0 | 6 | 5.6±0.2 | 113 | 1.0 | 6.0 | 3 | 3.2±0.1 | t=10.37, df=239, p<0.001* |
Cp10 | 166 | 1.0 | 15.0 | 7 | 6.9±0.2 | 161 | 2.0 | 10.0 | 4 | 3.7±0.1 | t=11.68, df=325, p<0.001* |
Cp20 | 78 | 1.0 | 15.0 | 8 | 7.0±0.3 | 73 | 2.0 | 8.0 | 4 | 3.7±0.2 | t=8.25, df=149, p<0.001* |
Cp30 | 134 | 2.0 | 10.0 | 6 | 5.9±0.2 | 118 | 1.0 | 6.0 | 4 | 3.8±0.1 | t=9.10, df=250, p<0.001* |
Ec10 | 186 | 1.0 | 15.0 | 7 | 6.9±0.2 | 178 | 2.0 | 10.0 | 4 | 3.8±0.1 | t=11.90, df=362, p<0.001* |
Ec20 | 127 | 1.0 | 15.0 | 7 | 7.1±0.3 | 122 | 2.0 | 10.0 | 4 | 3.9±0.2 | t=10.26, df=247, p<0.001* |
Ec30 | 170 | 1.0 | 15.0 | 7 | 6.9±0.2 | 162 | 2.0 | 10.0 | 4 | 3.7±0.1 | t=11.62, df=330, p<0.001* |
Td10 | 120 | 1.0 | 15.0 | 7 | 6.9±0.3 | 115 | 2.0 | 10.0 | 4 | 3.8±0.2 | t=9.88, df=233, p<0.001* |
Td20 | 173 | 1.0 | 15.0 | 7 | 6.9±0.2 | 165 | 2.0 | 10.0 | 4 | 3.7±0.1 | t=11.79, df=336, p<0.001* |
Td30 | 131 | 2.0 | 12.0 | 6 | 5.8±0.2 | 116 | 1.0 | 6.0 | 4 | 3.8±0.1 | t=8.66, df=245, p<0.001* |
Pooled plots | 1526 | 1.0 | 15.0 | 7 | 6.6±0.1 | 1431 | 1.0 | 10.0 | 4 | 3.7±0.0 | t=33.9, df=2955, p<0.001* |
ANOVA | F(10; 1,515)=5.467, p<0.001 * | F(10; 1,420)=1.675, p=0.082 ns | |||||||||
Pooled campaigns | |||||||||||
Untreated | 258 | 1.0 | 15.0 | 9 | 8.4±0.2 | 248 | 2.0 | 10.0 | 5 | 5.3±0.2 | t=11.90, df=504, p<0.001* |
Parastar | 284 | 2.0 | 10.0 | 6 | 5.8±0.1 | 254 | 1.0 | 6.0 | 3 | 3.5±0.1 | t=14.20, df=536, p<0.001* |
Cp10 | 318 | 1.0 | 15.0 | 7 | 7.0±0.2 | 308 | 2.0 | 10.0 | 4 | 3.7±0.1 | t=16.40, df=624, p<0.001* |
Cp20 | 271 | 1.0 | 20.0 | 8 | 7.4±0.2 | 251 | 1.0 | 10.0 | 4 | 4.5±0.2 | t=9.68, df=520, p<0.001* |
Cp30 | 325 | 1.0 | 15.0 | 6 | 6.5±0.2 | 304 | 1.0 | 10.0 | 4 | 3.7±0.1 | t=15.47, df=627, p<0.001* |
Ec10 | 397 | 1.0 | 20.0 | 8 | 7.4±0.2 | 381 | 2.0 | 11.0 | 4 | 4.3±0.1 | t=16.10, df=776, p<0.001* |
A. Duration (s) of the nectar collection | B. Duration (s) of the pollen collection | A vs. B | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
n | Min. | Max. | Median | Mean ± se | n | Min. | Max. | Median | Mean ± se | Student t-test | |
Ec20 | 325 | 1.0 | 20.0 | 8 | 8.2±0.2 | 312 | 2.0 | 10.0 | 4 | 4.8±0.1 | t=13.60, df=635, p<0.001* |
Ec30 | 365 | 1.0 | 15.0 | 7 | 6.9±0.2 | 349 | 2.0 | 10.0 | 4 | 3.7±0.1 | t=17.34, df=712, p<0.001* |
Td10 | 258 | 1.0 | 15.0 | 7 | 7.0±0.2 | 248 | 2.0 | 10.0 | 4 | 3.7±0.1 | t=14.74, df=504, p<0.001* |
Td20 | 326 | 1.0 | 15.0 | 7 | 7.6±0.2 | 303 | 1.0 | 10.0 | 4 | 4.7±0.1 | t=12.54, df=627, p<0.001* |
Td30 | 234 | 1.0 | 15.0 | 6 | 6.1±0.2 | 214 | 1.0 | 10.0 | 4 | 3.8±0.1 | t=11.75, df=446, p<0.001* |
Pooled plots | 3361 | 1.0 | 20.0 | 7 | 7.1±0.1 | 3172 | 1.0 | 11.0 | 4 | 4.2±0.0 | t=44.1, df=6531, p<0.001* |
ANOVA | F(10; 3,350)=18.156, p<0.001 * | F(10; 3,350)=23.356, p<0.001 * | |||||||||
C. Student-Nerwman-Keuls p-values for nectar collection: 2021 (upper diagonal matrix) and 2022 (diagonal lower matrix)
Parastar | Cp10 | Cp20 | Cp30 | Ec10 | Ec20 | Ec30 | Td10 | Td20 | Td30 | |
|---|---|---|---|---|---|---|---|---|---|---|
Untreated | 1x10-5 * | 2x10-5 * | 4x10-5 * | 3x10-5 * | 9x10-3* | 0.477 ns | 1x10-5 * | 3x10-5* | 0.031 * | 1x10-5* |
Parastar | - | 0.028 * | 2x10-4 * | 0.022 * | 3x10-5* | 1x10-5 * | 0.013 * | 0.034 * | 1x10-5* | 0.128 ns |
Cp10 | 7x10-4 * | - | 0.220 ns | 0.934 ns | 0.054 ns | 2x10-5 * | 0.979 ns | 0.914 ns | 0.008 * | 0.747 ns |
Cp20 | 0,009 * | 0,995 ns | - | 0.335 ns | 0.256 ns | 7x10-5 * | 0.415 ns | 0.391 ns | 0.085 ns | 0.173 ns |
Cp30 | 0,571 ns | 0,009 * | 0,078 ns | - | 0.036 * | 3x10-5 * | 0.966 ns | 0.818 ns | 0.003 * | 0.611 ns |
Ec10 | 6x10-4 * | 0,992 ns | 0,948 ns | 0,015 * | - | 0.002 * | 0.042 * | 0.084 ns | 0.271 ns | 0.015 * |
Ec20 | 8x10-4 * | 0,993 ns | 0,926 ns | 0,020 * | 0,963 ns | - | 3x10-5 * | 2x10-5 * | 0.046 * | 1x10-5 * |
Ec30 | 4x10-4 * | 0,977 ns | 0,998 ns | 0,003 * | 0,999 ns | 0,997 ns | - | 0.961 ns | 0.003 * | 0.360 ns |
Td10 | 0,003 * | 0,996 ns | 0,983 ns | 0,032 * | 0,918 ns | 0,984 ns | 0,999 ns | - | 0.012 * | 0.698 ns |
Td20 | 0,001 * | 0,999 ns | 0,768 ns | 0,024 * | 0,987 ns | 0,890 ns | 1.00 ns | 0,993 ns | - | 0.001* |
Td30 | 0,477 ns | 0,006 * | 0,050 ns | 0,768 ns | 0,008 * | 0,010 * | 0,004 * | 0,022 * | 0,012 * | |
Untreated | 3x10-4 * | 0,965 ns | 0,911 ns | 0,010 * | 0,911 ns | 0,715 ns | 0,978 ns | 0,953 ns | 0,838 ns | 0,005 * |
Pooled campaigns | ||||||||||
Untreated | 1x10-5 * | 2x10-5 * | 0,003 * | 1x10-5 * | 0,001 * | 0,671 ns | 3x10-5 * | 3x10-5 * | 0,006 * | 1x10-5 * |
Parastar | 5x10-5 * | 3x10-5 * | 0,008 * | 3x10-5 * | 1x10-5 * | 3x10-5 * | 1x10-4 * | 1x10-5 * | 0,168 ns | |
Cp10 | 0,115 ns | 0,273 ns | 0,121 ns | 2x10-5 * | 0,957 ns | 0,970 ns | 0,082 ns | 0,019 * | ||
Cp20 | 0,008 * | 0,825 ns | 0,004 * | 0,229 ns | 0,275 ns | 0,770 ns | 2x10-4 * | |||
Cp30 | 0,002 * | 3x10-5 * | 0,114 ns | 0,218 ns | 6x10-4 * | 0,158 ns | ||||
Ec10 | 0,002 * | 0,133 ns | 0,234 ns | 0,600 ns | 4x10-5 * | |||||
Ec20 | 3x10-5 * | 3x10-5 * | 0,005 * | 1x10-5 * | ||||||
Ec30 | 0,819 ns | 0,068 ns | 0,011* | |||||||
Td10 | 0,15 ns | 0,021 * | ||||||||
Td20 | 4x10-5 * | |||||||||
Parastar | Cp10 | Cp20 | Cp30 | Ec10 | Ec20 | Ec30 | Td10 | Td20 | Td30 | |
|---|---|---|---|---|---|---|---|---|---|---|
D. Student-Nerwman-Keuls p-values for pollen collection: 2021 (upper part matrix) and pooled years (lower part matrix) (Continued) | ||||||||||
Untreated | 2x10-5 * | 3x10-5 * | <0,001 * | 1x10-5 * | 2x10-5 * | 2x10-4 * | 1x10-5 * | 1x10-5 * | 0,020 * | 3x10-5 * |
Parastar | - | 0,879 ns | 8x10-5 * | 0,983 ns | 1x10-4 * | <0,001* | 0,994 ns | 0,992 ns | 2x10-5 * | 0,974 ns |
Cp10 | 0,673 ns | - | 5x10-5 * | 0,995 ns | 2x10-4 * | 2x10-5 * | 0,980 ns | 0,991 ns | 2x10-5 * | 0,933 ns |
Cp20 | 3x10-5 * | 7x10-5 * | - | 3x10-5 * | 0,555 ns | 0,009 * | 4x10-5 * | 1x10-4 * | 2x10-4 * | 5x10-4 * |
Cp30 | 0,470 ns | 0,986 ns | 9x10-5 * | - | 5x10-5 * | 3x10-5 * | 0,988 ns | 0,812 ns | 1x10-5 * | 0,965 ns |
Ec10 | 9x10-5 * | 0,002 * | 0,168 ns | 0,003 * | - | 0,003 * | 6x10-5 * | 5x10-4 * | 3x10-5 * | 0,002 * |
Ec20 | 1x10-5 * | 2x10-5 * | 0,129 ns | 3x10-5 * | 0,002 * | - | 1x10-5 * | 3x10-5 * | 0,115 ns | 2x10-5 * |
Ec30 | 0,279 ns | 0,991 ns | 5x10-5 * | 0,895 ns | 0,002 * | 1x10-5 * | - | 0,966 ns | 1x10-5 * | 0,993 ns |
Td10 | 0,621 ns | 0,955 ns | 2x10-4 * | 0,923 ns | 0,006 * | 3x10-5 * | 0,973 ns | - | 3x10-5 * | 0,972 ns |
Td20 | 1x10-5 * | 2x10-5 * | 0,311 ns | 3x10-5 * | 0,027 * | 0,336 ns | 3x10-5 * | 2x10-5 * | - | 3x10-5 * |
Td30 | 0,750 ns | 0,878 ns | 3x10-4 * | 0,991 ns | 0,003 * | 2x10-5 * | 0,993 ns | 0,979 ns | 1x10-5 * | |
Untreated | 1x10-5 * | 3x10-5 * | 6x10-5 * | 1x10-5 * | 2x10-5 * | 0,006 * | 1x10-5 * | 3x10-5 * | 8x10-4 * | 2x10-5 * |
Parastar | Cp10 | Cp20 | Cp30 | Ec10 | Ec20 | Ec30 | Td10 | Td20 | Td30 | |
|---|---|---|---|---|---|---|---|---|---|---|
A. Student-Newman-Keuls p-values: 6-7 a.m. in 2021 (upper diagonal matrix) and 8-9 a.m. in the pooled years (lower matrix) | ||||||||||
Untreated | p=0,811 ns | 1.00 ns | 0.705ns | 0,766 ns | 1.00 ns | 2x10-3 * | 0.933 ns | 0,814 ns | 0.968 ns | 1.00 ns |
Parastar | - | 1.00 ns | 0.419ns | 0.535 ns | 1.00 ns | 0,009 * | 0.443 ns | 0.629 ns | 0.718 ns | 1.00 ns |
Cp10 | 0.842 ns | - | 0,766ns | 0,814 ns | 1.00 ns | 9x10-4 * | 0,968 ns | 0,851ns | 0,984 ns | 1.00 ns |
Cp20 | 0.878 ns | 0.959 ns | - | 1.00 ns | 0.535 ns | 6x10-4 * | 0.718 ns | 1.00 ns | 0.443 ns | 0.639 ns |
Cp30 | 0.959 ns | 0.878ns | 0.863ns | - | 0,629ns | 7x10-4 * | 0.862 ns | 1.00 ns | 0.718 ns | 0.705 ns |
Ec10 | 0.863 ns | 0.981ns | 0.959ns | 0.816 ns | - | 6x10-3 * | 0.718 ns | 0.705 ns | 0.862 ns | 1.00 ns |
Ec20 | 0.019 * | 0.081ns | 0.074ns | 0.012 * | 0.032 * | - | 2x10-3 * | 8x10-4 * | 3x10-3 * | 4x10-3 * |
Ec30 | 0.784 ns | 0.805ns | 0.811ns | 1.00 ns | 0.771 ns | 0.010 * | - | 0.933 ns | 1.00 ns | 0.862ns |
Td10 | 0.921 ns | 1.00 ns | 0.784ns | 0.925 ns | 0.946 ns | 0.059ns | 0.878 ns | - | 0.862ns | 0.966 ns |
Td20 | 0.959 ns | 0.584ns | 0.842ns | 0.981 ns | 0.878 ns | 0.026 * | 0.946 ns | 0.846 ns | - | 0.933 ns |
Td30 | 0.784 ns | 0.846ns | 0.921ns | 0.946 ns | 0.925 ns | 0.033 * | 0.846 ns | 0.946 ns | 1.00 ns | - |
Untreated | p=0,811 ns | 0,946ns | 0,784ns | 0.771 ns | 1.00 ns | 0.079 ns | 0.716 ns | 0.846 ns | 0.805 ns | 0.878 ns |
B. One-way ANOVA in each plot | ||||||||||
2021 (132 essays) | 2022 (132 essays) | Pooled campaigns (264 essays) | ||||||||
Plots | ANOVA | p-value | ANOVA | p-value | ANOVA | p-value | ||||
Untreated: | F(3; 8)=1.600 | 0.264 ns | F(3; 8)=4.083 | 0.050 ns | F(3; 20)=4.574 | 0.014 * | ||||
Parastar | F(3; 8)=4.267 | 0.045 * | F(3; 8)=3.074 | 0.091 ns | F(3; 20)=5.044 | 9.2x10-3 * | ||||
Cp10 | F(3; 8)=3.963 | 0.053 ns | F(3; 8)=3.852 | 0.056 ns | F(3; 20)=11.229 | 9.0x10-5 * | ||||
Cp20 | F(3; 8)=4.458 | 0.040 * | F(3; 8)=2.095 | 0.179 ns | F(3; 20)=8.049 | 7.0x10-4 * | ||||
Cp30 | F(3; 8)=1.267 | 0.349 ns | F(3; 8)=3.278 | 0.080 ns | F(3; 20)=6.162 | 2.9x10-3 * | ||||
Ec10 | F(3; 8)=3.769 | 0.059 ns | F(3; 8)=4.222 | 0.046 * | F(3; 20)=9.317 | 4.6x10-4 * | ||||
Ec20 | F(3; 8)=6.952 | 0.013 * | F(3; 8)=7.606 | 9.9x10-3 * | F(3; 20)=9.277 | 4.8x10-4 * | ||||
Ec30 | F(3; 8)=1.667 | 0.250 ns | F(3; 8)=3.571 | 0.067 ns | F(3; 20)=5.630 | 5.8x10-3 * | ||||
Td10 | F(3; 8)=3.619 | 0.065 ns | F(3; 8)=4.400 | 0.042 * | F(3; 20)=12.179 | 5.0x10-5 * | ||||
Td20 | F(3; 8)=1.296 | 0.341 ns | F(3; 8)=1.556 | 0.274 ns | F(3; 20)=3.429 | 0.037 * | ||||
Td30 | F(3; 8)=7.667 | 9.7x10-3 * | F(3; 8)=4.714 | 0.035 * | F(3; 20)=11.609 | 1.3x10-4 * | ||||
Global | F(3; 128)=27.67 | <0.001* | F(3; 128)=41.74 | <0.001* | F(3; 260)=60.84 | <0.001* | ||||
C. Between time periods in each category of plot: Student-Newman-Keuls test p-values
Parastar | Cp10 | Cp20 | Cp30 | Ec10 | Ec20 | Ec30 | Td10 | Td20 | Td30 | |
|---|---|---|---|---|---|---|---|---|---|---|
2021 campaign | 2022 campaign | |||||||||
Parastar | Cp20 | Ec20 | Td30 | Pooled | Ec10 | Ec20 | Td10 | Td30 | Pooled | |
6-7 vs. 8-9 a.m. | 1.00 ns | 0.085 ns | 0.290 ns | 0.545 ns | 5x10-5* | 0.076ns | 0.012 * | 0.056 ns | 0,065 ns | <0.001 * |
6-7 vs. 10-11 a.m. | 0.081 ns | 0.347 ns | 0.096 ns | 0.242 ns | 0.011* | 0.438ns | 0.681 ns | 1.00 ns | 1,00 ns | 0.468 ns |
6-7 vs. 12-13 p.m. | 0.129 ns | 0.347 ns | 0.039 * | 0.013 * | 2x10-5* | 0.473ns | 0.681 ns | 0.413ns | 0,299 ns | 0.036 * |
8-9 vs. 10-11 a.m. | 0.035 * | 0.172 ns | 0.039 * | 0.201 ns | 2x10-5* | 0.050ns | 9x10-3* | 0.124 ns | 0,143 ns | 2.0x10-5* |
8-9 vs. 12-13 p.m. | 0.081 ns | 0.033 * | 0.014 * | 0.010 * | <0,001* | 0.046 * | 0.012 * | 0.030 * | 0,0238 * | <0.001 * |
10-11 vs. 12-13 p.m. | 1.00 ns | 0.174 ns | 0.290 ns | 0.035 * | 7x10-3* | 0.694ns | 0.683ns | 0.217 ns | 0,148 ns | 0.082 ns |
Pooled campaigns | ||||||||||
6-7 vs. 8-9 a.m. | 0,015 * | 0,083 ns | 9x10-3* | 0,001* | 0,004 * | 0,023 * | 9x10-3 * | 0,253 ns | 6x10-4 * | 0,106 ns |
6-7 vs. 10-11 a.m. | 1,00 ns | 0,187 ns | 0,230ns | 0,356ns | 0,691ns | 0,139ns | 0,348 ns | 0,253 ns | 1,00 ns | 0,382 ns |
6-7 vs. 12-13 p.m. | 0,708 ns | 0,286 ns | 0,024 * | 0,757ns | 0,915ns | 0,057ns | 0,102 ns | 0,032 * | 0,183 ns | 0,506 ns |
8-9 vs. 10-11 a.m. | 0,038 * | 0,008 * | 2x10-3* | 0,005* | 0,004 * | 0,002 * | 3x10-3 * | 0,071 ns | 0,002 * | 0,030 * |
8-9 vs. 12-13 p.m. | 0,012 * | 0,022 * | 2x10-4* | 0,001* | 0,007 * | 6x10-4 * | 5x10-4 * | 4x10-3* | 2x10-4 * | 0,069 ns |
10-11vs. 12-13 p.m. | 0,434 ns | 0,474 ns | 0,090ns | 0,434ns | 1.00 ns | 0,366ns | 0,243 ns | 0,133 ns | 0,081 ns | 0,506 ns |
Td30 | Pooled | |||||||||
6-7vs. 8-9 a.m. | 0,051 ns | <0,001 * | ||||||||
6-7vs. 10-11 a.m. | 0,416 ns | 0,023 * | ||||||||
6-7vs. 12-13 p.m. | 0,004 * | 2x10-5 * | ||||||||
8-9vs. 10-11 a.m. | 0,023 * | 2x10-5 * | ||||||||
8-9vs. 12-13 p.m. | 2x10-4 * | <0,001* | ||||||||
10-11vs. 12-13 p.m. | 0,009 * | 0,002 * | ||||||||
Ax | Fx | V1000 =(Ax/Fx)*1000 | |||||
|---|---|---|---|---|---|---|---|
Essays | Min.-Max. | Mean ± se | Min.-Max. | Mean ± se | Min.-Max. | Mean ± se | |
A. 2021 campaign | |||||||
Untreated plots | 362 | 1-9 | 3±0 | 5-689 | 184 ± 10 | 1-800 | 31 ± 3 |
Parastar | 177 | 1-9 | 3±0 | 5-720 | 273 ± 14 | 2-400 | 17 ± 3 |
Cp10 | 215 | 1-9 | 3±0 | 5-689 | 260 ± 13 | 1-800 | 24 ± 4 |
Cp20 | 229 | 1-9 | 3±0 | 5-689 | 216 ± 12 | 2-800 | 29 ± 4 |
Cp30 | 214 | 1-9 | 3±0 | 5-689 | 256 ± 13 | 1-400 | 19 ± 2 |
Ec10 | 225 | 1-9 | 3±0 | 5-689 | 257 ± 12 | 1-800 | 23 ± 4 |
Ec20 | 324 | 1-9 | 3±0 | 5-800 | 201 ± 10 | 1-800 | 31 ± 3 |
Ec30 | 205 | 1-9 | 3±0 | 5-700 | 260 ± 13 | 1-400 | 18 ± 2 |
Td10 | 185 | 1-9 | 3±0 | 5-689 | 289 ± 14 | 1-800 | 23 ± 5 |
Td20 | 316 | 1-9 | 3±0 | 5-689 | 199 ± 10 | 1-800 | 28 ± 3 |
Td30 | 204 | 1-9 | 3±0 | 5-689 | 269 ± 13 | 1-400 | 19 ± 2 |
Global | 2,656 | 1-9 | 3±0 | 5-800 | 234 ± 4 | 1-800 | 25 ± 1 |
B. 2022 campaign | |||||||
Untreated plots | 388 | 1-9 | 3±0 | 5-689 | 193 ± 9 | 1-400 | 26 ± 2 |
Parastar | 191 | 1-9 | 3±0 | 24-800 | 269 ± 14 | 1-179 | 16 ±1 |
Cp10 | 246 | 1-9 | 3±0 | 5-689 | 293 ± 12 | 1-800 | 19 ± 3 |
Cp20 | 332 | 1-9 | 3±0 | 5-689 | 223 ± 10 | 1-800 | 24 ± 3 |
Cp30 | 232 | 1-9 | 3±0 | 5-800 | 295 ± 14 | 1-800 | 18 ± 4 |
Ec10 | 247 | 1-9 | 3±0 | 5-700 | 274 ± 12 | 1-800 | 22 ± 4 |
Ec20 | 356 | 1-9 | 3±0 | 5-689 | 218 ± 10 | 1-800 | 24 ± 3 |
Ec30 | 247 | 1-9 | 3±0 | 5-800 | 298 ± 13 | 1-800 | 18 ± 3 |
Td10 | 228 | 1-9 | 3±0 | 5-689 | 239 ± 13 | 1-400 | 22 ± 2 |
Td20 | 241 | 1-9 | 3±0 | 5-689 | 239 ± 12 | 1-800 | 24 ± 4 |
Td30 | 218 | 1-9 | 3±0 | 5-689 | 249 ± 13 | 1-400 | 20 ± 2 |
Global | 2,939 | 1-9 | 3±0 | 5-800 | 248 ± 4 | 1-800 | 22 ± 1 |
C. Pooled campaigns | |||||||
Untreated plots | 750 | 1-9 | 3±0 | 5-689 | 189 ± 7 | 1-800 | 28 ± 2 |
Parastar | 368 | 1-9 | 3±0 | 5-800 | 271 ± 10 | 1-400 | 17 ± 1 |
Cp10 | 461 | 1-9 | 3±0 | 5-689 | 277 ± 9 | 1-800 | 22 ± 3 |
Ax | Fx | V1000 =(Ax/Fx)*1000 | |||||
|---|---|---|---|---|---|---|---|
Essays | Min.-Max. | Mean ± se | Min.-Max. | Mean ± se | Min.-Max. | Mean ± se | |
C. Pooled campaigns (Continued) | |||||||
Cp20 | 561 | 1-9 | 3±0 | 5-689 | 220 ± 8 | 1-800 | 26 ± 2 |
Cp30 | 446 | 1-9 | 3±0 | 5-800 | 276 ± 9 | 1-800 | 19 ± 2 |
Ec10 | 472 | 1-9 | 3±0 | 5-700 | 266 ± 9 | 1-800 | 22 ± 3 |
Ec20 | 680 | 1-9 | 3±0 | 5-800 | 210 ± 7 | 1-800 | 27 ± 2 |
Ec30 | 452 | 1-9 | 3±0 | 5-800 | 281 ± 9 | 1-800 | 18 ± 3 |
Td10 | 413 | 1-9 | 3±0 | 5-689 | 261 ± 10 | 1-800 | 22 ± 2 |
Td20 | 557 | 1-9 | 3±0 | 5-689 | 216 ± 8 | 1-800 | 26 ± 2 |
Td30 | 422 | 1-9 | 3±0 | 5-689 | 259 ± 9 | 1-400 | 19 ± 2 |
Global | 5,595 | 1-9 | 3±0 | 5-800 | 242 ± 3 | 1-800 | 23 ± 1 |
Visited flowers Fi | Foraging duration (seconds): di | Vb=(Fi/di)*60 | |||||
|---|---|---|---|---|---|---|---|
Essays | Min.-Max. | Mean ± se | Min.-Max. | Mean ± se | Min.-Max. | Mean ± se | |
A. 2021 campaign | |||||||
Untreated plots | 221 | 1-3 | 1±0 | 2.0-60.0 | 13.5±0.6 | 1-60 | 9±1 |
Parastar | 116 | 1-3 | 1±0 | 3.0-60.0 | 22.1±1.3 | 1-20 | 6±0 |
Cp10 | 140 | 1-3 | 1±0 | 3.0-60.0 | 23.5±1.3 | 1-20 | 5±0 |
Cp20 | 156 | 1-3 | 1±0 | 3.0-60.0 | 18.9±1.1 | 1-30 | 7±0 |
Cp30 | 102 | 1-3 | 1±0 | 3.0-56.0 | 17.6±1.1 | 1-24 | 6±0 |
Ec10 | 150 | 1-3 | 1±0 | 3.0-60.0 | 20.2±1.3 | 1-40 | 7±1 |
Ec20 | 201 | 1-3 | 1±0 | 1.0-60.0 | 17.8±1.0 | 1-40 | 9±1 |
Ec30 | 120 | 1-3 | 1±0 | 3.0-60.0 | 20.3±1.3 | 1-40 | 7±0 |
Td10 | 145 | 1-3 | 1±0 | 3.0-60.0 | 23.3±1.3 | 1-20 | 5±0 |
Td20 | 160 | 1-3 | 1±0 | 3.0-60.0 | 18.8±1.0 | 1-30 | 7±0 |
Td30 | 111 | 1-3 | 1±0 | 3.0-60.0 | 22.1±1.4 | 1-30 | 6±0 |
Global | 1622 | 1-3 | 1±0 | 1.0-60.0 | 19.4±0.3 | 1-120 | 7±0 |
ANOVA | F(10; 1611)=0.043, p=1.00 ns | F(10; 1611)=8.130, p<0.001 * | F(10; 681)=4.23, p=1x10-5 * | ||||
B. 2022 campaign | |||||||
Untreated plots | 230 | 1-3 | 1±0 | 3.0-60.0 | 18.4±1.0 | 1-24 | 5±0 |
Parastar | 76 | 1-3 | 1±0 | 3.0-60.0 | 22.2±1.6 | 1-20 | 5±0 |
Cp10 | 124 | 1-3 | 1±0 | 3.0-60.0 | 19.1±1.2 | 1-24 | 7±0 |
Cp20 | 187 | 1-2 | 1±0 | 3.0-60.0 | 19.3±1.1 | 1-30 | 7±0 |
Cp30 | 103 | 1-2 | 1±0 | 3.0-60.0 | 20.9±1.4 | 1-30 | 6±0 |
Ec10 | 122 | 1-3 | 1±0 | 3.0-60.0 | 19.2±1.2 | 1-60 | 7±0 |
Ec20 | 214 | 1-2 | 1±0 | 3.0-60.0 | 19.3±1.0 | 1-24 | 7±0 |
Ec30 | 108 | 1-2 | 1±0 | 3.0-60.0 | 20.4±1.4 | 1-30 | 6±0 |
Td10 | 134 | 1-3 | 1±0 | 3.0-60.0 | 20.6±1.2 | 1-60 | 6±1 |
Td20 | 166 | 1-2 | 1±0 | 3.0-60.0 | 19.4±1.1 | 1-24 | 7±0 |
Td30 | 105 | 1-2 | 1±0 | 3.0-60.0 | 22.9±1.6 | 1-30 | 6±0 |
Global | 1569 | 1-3 | 1±0 | 3.0-60.0 | 19.8±0.4 | 1-60 | 7±0 |
ANOVA | F(10; 1558)=0.100, p=1.00 ns | F(10; 1558)=1.164, p=0.311 ns | F(10; 627)=0.40, p=0.948 ns | ||||
C. Pooled campaigns | |||||||
Untreated plots | 451 | 1-3 | 1±0 | 2.0-60.0 | 16.0±0.6 | 1-60 | 7±0 |
Visited flowers Fi | Foraging duration (seconds): di | Vb=(Fi/di)*60 | |||||
|---|---|---|---|---|---|---|---|
Essays | Min.-Max. | Mean ± se | Min.-Max. | Mean ± se | Min.-Max. | Mean ± se | |
Parastar | 192 | 1-3 | 1±0 | 3.0-60.0 | 22.1±1.0 | 1-20 | 6±0 |
Cp10 | 264 | 1-3 | 1±0 | 3.0-60.0 | 21.5±0.9 | 1-24 | 6±0 |
Cp20 | 343 | 1-3 | 1±0 | 2.0-60.0 | 19.1±0.8 | 1-30 | 7±0 |
Cp30 | 205 | 1-3 | 1±0 | 3.0-60.0 | 19.3±0.9 | 1-30 | 6±0 |
Ec10 | 272 | 1-3 | 1±0 | 3.0-60.0 | 19.8±0.9 | 1-40 | 7±0 |
Ec20 | 415 | 1-3 | 1±0 | 1.0-60.0 | 18.6±0.7 | 1-120 | 8±0 |
Ec30 | 228 | 1-3 | 1±0 | 2.0-60.0 | 20.3±0.9 | 1-30 | 6±0 |
Td10 | 279 | 1-3 | 1±0 | 2.0-60.0 | 22.0±0.9 | 1-60 | 6±0 |
Td20 | 326 | 1-3 | 1±0 | 3.0-60.0 | 19.1±0.8 | 1-30 | 7±0 |
Td30 | 216 | 1-3 | 1±0 | 3.0-60.0 | 22.5±1.1 | 1-30 | 6±0 |
Global | 3191 | 1-3 | 1±0 | 1.0-60.0 | 19.6±0.2 | 1-120 | 7±0 |
ANOVA | F(10; 3180)=0.096, p=1.00 ns | F(10; 3180)=6.23, p<0.001 * | F(10; 1319)=3.61, p=1x10-4 * | ||||
D. 2021 campaign vs. 2022 campaign: Student t-test
Visited flowers Fi | Duration (seconds): di | Foraging speed Vb | |
|---|---|---|---|
Untreated plots | t= 0.258; df=449; p=0.796 ns | t=-4.266; df=449; p=2x10-5 * | t=3.473; df=449; p=8x10-4 * |
Parastar | t=-0.156; df=190; p=0.876 ns | t=-0.024; df=190; p=0.981 ns | t=0.340; df=190; p=0.734 ns |
Cp10 | t=0.018; df=262; p=0.986 ns | t=2.457; df=262; p=0.015 * | t=-2.200; df=262; p=0.029 * |
Cp20 | t=-0.189; df=341; p=0.850 ns | t=-0.305; df=341; p=0.760 ns | t=0.019; df=341; p=0.985 ns |
Cp30 | t=0.328; df=203; p=0.743 ns | t=-1.814; df=203; p=0.071 ns | t=-0.573; df=203; p=0.568 ns |
Ec10 | t=-0.225; df=270; p=0.822 ns | t=-0.590; df=270; p=0.555 ns | t=-0.461; df=270; p=0.645 ns |
Ec20 | t=-0.144; df=413; p=0.885 ns | t=-1.021; df=413; p=0.308 ns | t=2.333; df=413; p=0.020 * |
Ec30 | t=-0.373; df=226; p=0.710 ns | t=-0.043; df=226; p=0.966 ns | t=-0.514; df=226; p=0.607 ns |
Td10 | t=-0.154; df=277; p=0.878 ns | t=1.546; df=277; p=0.123 ns | t=1.558; df=277; p=0.120 ns |
Td20 | t=0.492; df=324; p=0.623 ns | t=-0.374; df=324; p=0.708 ns | t=0.034; df=324; p=0.973 ns |
Td30 | t=-0.476; df=665; p=0.634 ns | t=-3.219; df=665; p=1.4x10-3 * | t=3.082; df=665; p=2.1x10-3 * |
Global | t=0.673; df=3189; p=0.501 ns | t=-0.825; df=3189; p=0.409 ns | t=2.008; df=3189; p=0.045 * |
E. Student-Newman-Keuls p-values for foraging duration 2021 campaign (upper part matrix) and 2022 (lower matrix)
Parastar | Cp10 | Cp20 | Cp30 | Ec10 | Ec20 | Ec30 | Td10 | Td20 | Td30 | |
|---|---|---|---|---|---|---|---|---|---|---|
Untreated | 3x10-5 * | 1x10-5 * | 0.002 * | 0.012 * | 6x10-5 * | 0.004 * | 3x10-4 * | 1x10-5 * | 0.001 * | 1x10-5 * |
Parastar | - | 0.841ns | 0.208 ns | 0.188 ns | 0.500 ns | 0.073 ns | 0.304 ns | 0.757 ns | 0.272 ns | 0.994 ns |
Cp10 | 0.868 ns | - | 0.050 ns | 0.030 * | 0.305 ns | 0.004 * | 0.310 ns | 0.896 ns | 0.056 ns | 0.697 ns |
Cp20 | 0.202 ns | 0.243ns | - | 0.894 ns | 0.381 ns | 0.749 ns | 0.662 ns | 0.052 ns | 0.976 ns | 0.300 ns |
Cp30 | 0.319 ns | 0.330ns | 0.905ns | - | 0.574 ns | 0.916 ns | 0.697 ns | 0.034 * | 0.773 ns | 0.238 ns |
Ec10 | 0.375 ns | 0.338ns | 0.836ns | 0.700 ns | - | 0.354 ns | 0.968 ns | 0.290 ns | 0.633 ns | 0.679 ns |
Parastar | Cp10 | Cp20 | Cp30 | Ec10 | Ec20 | Ec30 | Td10 | Td20 | Td30 | |
|---|---|---|---|---|---|---|---|---|---|---|
E. Student-Newman-Keuls p-values for foraging duration 2021 campaign (upper part matrix) and 2022 (lower matrix) (Continued) | ||||||||||
Ec20 | 0.081 ns | 0.114ns | 0.845ns | 0.933 ns | 0.802 ns | - | 0.510 ns | 0.005 * | 0.486 ns | 0.103 ns |
Ec30 | 0.552 ns | 0.371ns | 0.741ns | 0.709 ns | 0.652 ns | 0.638 ns | - | 0.272 ns | 0.806 ns | 0.561 ns |
Td10 | 0.930 ns | 0.644ns | 0.104 ns | 0.203 ns | 0.232 ns | 0.030 * | 0.367 ns | - | 0.060 ns | 0.488 ns |
Td20 | 0.253 ns | 0.324ns | 0.994 ns | 0.991 ns | 0.941 ns | 0.592 ns | 0.850 ns | 0.142 ns | - | 0.359 ns |
Td30 | 0.780 ns | 0.843ns | 0.095 ns | 0.204 ns | 0.256 ns | 0.025 * | 0.466 ns | 0.917 ns | 0.121 ns | - |
Untreated | 3x10-5 * | 4x10-5 * | 0.011 * | 0.046 * | 0.006 * | 0.008 * | 0.003 * | 1x10-5 * | 0.007 * | 2x10-5 * |
F. Student-Newman-Keuls p-values for foraging speed 2021 campaign (upper diagonal matrix) and 2022 (lower matrix) | ||||||||||
Untreated | 0.007* | 0.002 * | 0.077ns | 0.040* | 0.058ns | 0.810ns | 0.085ns | 0.002 * | 0.043 * | 0.055 ns |
Parastar | - | 0.936ns | 0.710ns | 0.588ns | 0.723ns | 0.012 * | 0.821ns | 0.995ns | 0.701ns | 0.776 ns |
Cp10 | 0.597ns | - | 0.688ns | 0.793ns | 0.683ns | 0.005 * | 0.835ns | 0.986ns | 0.734ns | 0.858 ns |
Cp20 | 0.527ns | 0.729ns | - | 0.912ns | 0.909ns | 0.076ns | 0.767ns | 0.735ns | 0.913ns | 0.897 ns |
Cp30 | 0.917ns | 0.988ns | 0.688ns | - | 0.929ns | 0.058ns | 0.952ns | 0.908ns | 0.873ns | 0.887 ns |
Ec10 | 0.561ns | 0.764ns | 0.907ns | 0.748ns | - | 0.040 * | 0.914ns | 0.721ns | 0.953ns | 0.929 ns |
Ec20 | 0.020 * | 0.036 * | 0.200ns | 0.059ns | 0.132ns | - | 0.094ns | 0.005 * | 0.058ns | 0.073 ns |
Ec30 | 0.843ns | 0.950ns | 0.751ns | 0.842ns | 0.853ns | 0.141ns | - | 0.882ns | 0.939ns | 0.922 ns |
Td10 | 0.807ns | 0.921ns | 0.703ns | 0.958ns | 0.748ns | 0.036 * | 0.919ns | - | 0.806ns | 0.926 ns |
Td20 | 0.807ns | 0.940ns | 0.524ns | 0.884ns | 0.747ns | 0.078ns | 0.892ns | 0.921ns | - | 0.734 ns |
Td30 | 0.872ns | 0.961 ns | 0.788ns | 0.738ns | 0.854 ns | 0.129ns | 0.830 ns | 0.915ns | 0.930 ns | - |
Untreated | 0.009* | 0.015 * | 0.172ns | 0.031 * | 0.163ns | 0.734ns | 0.094ns | 0.016 * | 0.050ns | 0.080 ns |
ANOVA | Analysis of Variance |
Ah. craccivora | Aphis craccivora Koch, 1854 |
Am. calens | Amegilla calens (Le Peletier, 1841) |
Ap. mellifera | Apis mellifera Linnaeus, 1758 |
Ca. procera | Calotropis procera (Aiton) W. T. Aiton, 1811 |
E. camaldulensis | Eucalyptus camaldulensis Dehnh., 1832 |
FAO | Food and Agriculture Organization |
FAOSTAT | Food and Agriculture Organization Statistics |
H. misippus | Hypolimnas misippus (Linnaeus, 1764) |
MINADER | Ministry of Agriculture and Rural Development (Cameroon) |
MINEF | Ministry of Environment and Forestry (Cameroon) |
V. unguiculata | Vigna unguiculata (L.) Walp., 1843 |
Ti. diversifolia | Tithonia diversifolia (Hemsl.) A. Gray, 1883 |
X. caffra | Xylocopa caffra (L. 1767) |
X. erythrina | Xylocopa erythrina Gribodo 1894 |
X. imitator | Xylocopa imitator Smith, 1854 |
X. inconstans | Xylocopa inconstans Smith F. 1874 |
X. nigrita | Xylocopa nigrita (Fabricius 1775) |
X. olivacea | Xylocopa. olivacea (Fabricius 1778) |
| [1] | Abebe, B. K., Alemayehu, M. T. A review of the nutritional use of cowpea (Vigna unguiculata L. Walp) for human and animal diets. Journal of Agriculture and Food Research, 202210, 100383. |
| [2] | Omoigui, L. O., Kamara, A. Y., Batieno, J., Iorlamen, T., Kouyate, Z., Yirzagla, J., Garba, U., Diallo, S. Guide sur la production de niébé en Afrique de l’Ouest. Ibadan, Nigeria: International Institute of Tropical Agriculture (IITA); 2018, pp. 1-65. |
| [3] | Fery, R. L. The genetics of cowpea: a review of the world literature. In Cowpea research, production and utilization, Singh, S. R., Rachie, K. O., eds., Wiley, New York, United States, 1985, pp. 25-62. |
| [4] | Brink, M., Belay, G. Plant Resources of Tropical Africa 1. Cereals and Pulses. Wageningen, Netherlands: PROTA Foundation, Backhuys Puplisher, Netherlands/CTA; 2006, 1-328. |
| [5] | Horn, L., Shimelis, H. Production constraints and breeding approaches for cowpea improvement for drought prone agro-ecologies in Sub-Saharan Africa. Annals of Agricultural Sciences. 2020, 65, 83-91. |
| [6] | Owade, J. O., Abong, G., Okoth, M., Mwang’ombe, A. W. A review of the contribution of cowpea leaves to food and nutrition security in East Africa. Food Science & Nutrition. 2020, 8, 36-47. |
| [7] | Singh, B. B., Ajeigbe, H. A., Tarawali, S. A., Fernandez-Rivera, S., Abubakar, M. Improving the production and utilization of cowpea as food and fodder. Field Crops Research. 2003, 84(s 1-2), 169-177. |
| [8] |
Amougou, P., Breuil, F., 2018. Le niébé, une légumineuse contre la malnutrition. Développée par l'IRAD au Cameroun. Available from:
https://www.mediaterre.org/afrique-centrale/actu,20181112153028.html (accessed on 19 Mai 2023). |
| [9] | Food and Agriculture Organization. FAO: Statistical Yearbook 2022. Rome, Italy, 2022, pp. 1-382. |
| [10] |
Food and Agriculture Organization Statistics. FAOSTAT: Cowpeas, dry. Available from:
http://www.fao.org/faostat/en/#data/QC (accessed on August 8, 2021). |
| [11] | Adamou, M., Kosini, D., Tchoubou-Salé, A., Massah, O. D., Tchocgnia, T. F. C., Mohammadou, M., Youssoufa, O., Nukenine, E. N. Impact of aqueous extracts of Cassia occidentalis, Eucalyptus camaldulensis and Hyptis suaveolens on the entomofauna and the seed yield of Gossypium hirsutum at Bokle (Garoua, Cameroon). Heliyon. 2022, 8(10), e 10937. |
| [12] | Son, D., Somda, I., Legreve, A., Schiffers, B. Pratiques phytosanitaires des producteurs de tomates du Burkina Faso et risques pour la santé et l’environnement. Cahier Agriculture. 2017, 26(2), 25005-25008. |
| [13] |
Ministry of Agriculture and Rural Development (Cameroon). MINADER: Liste des pesticides homologués au Cameroun au 18 Avril 2019. Liste réservée au grand public. Ministère de l’Agriculture et de Développement Rural. Commission Nationale d’Homologation des Produits Phytosanitaires et de Certification des Appareils de Traitement (CNHPPCZT), Yaoundé, Cameroun, 2019. Available from
https://drcq-minader.org/docs/Liste_Pesticides_Homologues_042019.pdf |
| [14] | Mohammadou, M., Adamou, M., Taïmanga, Kosini, D., Kenne, M. Seed Yield Improvement in Vigna unguiculata (L.) (Fabaceae): Efficiency of Pollinators and Impact of Aqueous Leaf Extract of Three Plant Species in North Cameroon. Asian Journal of Crop Science. 2023a, 8(3), 146-172. |
| [15] | McGregor, S. E. Insect pollination of cultivated crop plants. Agricultural Research Service. Washington: United States Department of Agriculture, Agric. Handb. 496; 1976, pp. 1-411. |
| [16] | Faegri, K., Pijl, L. V. D. The principle of pollination ecology. 3rd revised edition; Oxford: Pergamon Press; 1979, pp. 1-244. |
| [17] | Segeren, P., Mulder, V., Beetsma, J., Sommeijer, R. Apiculture sous les tropiques. Agrodok 32, 5ème edition; Wageningen: Agromisa; 1996, pp. 1-88. |
| [18] | Tchuenguem Fohouo, F.-N., Ngakou, A., Kengni, B. S. Pollination and yield responses of cowpea (Vigna unguiculata L. Walp.) to the foraging activity of Apis mellifera adansonii (Hymenoptera: Apidae) at Ngaoundéré (Cameroon). African Journal of Biotechnology. 2009, 8(9), 1988-1996. |
| [19] | Djonwangwe, D, Pando, J. B., Balle, K., Mbonoma, B., Tchuenguem Fohouo, F.-N., Messi, J. Impact des activités de butinage de Xylocopa inconstans Smith F. 1874 (Hymenoptera: Apidae) et Megachile eurymera Smith 1864 (Hymenoptera: Megachilidae) sur la pollinisation et les rendements fruiter et grainier de Vigna unguiculata (L.) Walp. 1843 (Fabaceae) à Maroua, Extrême-Nord, Cameroun. Afrique Science. 2017, 13(5), 1-17. |
| [20] | Adamou, M., Nepide, N. C., Mazi, S., Yatahaï, C. M. Impact of the pollinating activity of Apis mellifera (Hymenoptera: Apidae) on pod and seed yields of Vigna unguiculata (Fabaceae) variety BR1 in Djoumassi (North Cameroon). Cameroon Journal of Biological and Biochemical Sciences. 2020, 28(2), 146-159. |
| [21] | Mazi, S., Kingha Tekombo, B. M., Adamou, M., Yatahaï, C. M. Impact of the foraging activity of Apis mellifera (Hymenoptera: Apidae) on increasing yields of Gossypium hirsutum (Malvaceae) in Djoumassi (Garoua, Cameroon). GSC Biological and Pharmaceutical Sciences. 2020, 12(1), 255-266. |
| [22] | Desquesne, P. H.. Apiculture tropicale en Afrique de l’Ouest, L’abeille de France. 1996, 31-132. |
| [23] | Mbianda, P. A., Douka, C., Dounia, Eloundou, C. E., Tchuenguem Fohouo. F.-N. Pollination efficiency of Apis mellifera L. (Hymenoptera: Apidae) on flowers of Vigna unguiculata (L.) Walp. (Fabaceae) at Bilone (Obala, Cameroon). International Jopurnal of Bioscience. 2019, 14 (1): 1-11. |
| [24] | Tchekote, H., Nguiffeu Tajouo, E. L., Nguedia Melachio, M., Siyapdje, E. C., Mbeng, E. Farmers’ Accessibility to Pesticides and Generalization of Farming Practices besides the Legal Framework in Northern Bafou, in the Bamboutos Mountains (West Cameroon). Sustainability in Environment. 2019, 4(1), 41-56. |
| [25] | Yang, E., Chuang, Y., Chen, Y., Chang, L. Abnormal foraging behavior induced by sub-lethal dosage of imidacloprid in the honey bee (Hymenoptera: Apidae). Journal of Economic Entomology. 2008, 101: 1743-1748. |
| [26] | Sande, D., Mullen, J., Wetzstein, M., Houston, J. Environmental impacts from pesticide use: a case study of soil fumigation in Florida tomato production. International Journal of Environmental Research and Public Health. 2011, 8(12), 4649-4661. |
| [27] | Ndakidemi, B., Mtei, K., Ndakidemi, P. Impacts of Synthetic and Botanical Pesticides on Beneficial Insects. Agricultural Science. 2016, 7(6), 364-372. |
| [28] | Sonchieu, J., Ngassoum, M. B., Nantia, Akono, E., Laxman, P. S. Pesticide Applications on Some Vegetables Cultivated and Health Implications in Santa, North-West Cameroon. SSRG. International Journal of Agriculture and Environmental Research. 2018, 4(2), 39-46. |
| [29] | Galani, Y. J. H., Houbraken, M., Wumbei, A., Djeugap, J. F., Fotio, D., Gong, Y. Y., Spanoghe, P. Contamination of Foods from Cameroon with Residues of 20 Halogenated Pesticides, and Health Risk of Adult Human Dietary Exposure. International Journal of Environmental Research and Public Health. 2021, 18(9), 5043. |
| [30] | Sopkoutie, N. G. K., Abdulai A. N., Tarla, D. N., Galani, Y. J. H., Djeugap F. J., Ekengoue C. M., Tabang, W. M., Nya, E., Payne, V. K. Phytosanitary practices and evaluation of 17 pesticides residues in tomatoes fruits produced in Foumbot district Western Highland-Cameroon. European Scientific Journal. 2021, 17(1), 1. |
| [31] | Smith, C. J., Perfetti, T. A. A comparison of the persistence, toxicity, and exposure to high-volume natural plant-derived and synthetic pesticides. Toxicology Research and Application. 2020, 4, 1–15. |
| [32] | Thapa1, R., Nainabasti, A., Lamsal, A., Malla, S., Thapa, B., Subedi, Y., Ghimire, S. Pesticide Persistence in Agriculture and its hazardous effects on Environmental. International Journal of Applied Sciences and Biotechnology. 2022, 10(2), 75-83. |
| [33] | Gupta, N., Sharma, N. & Ramniwas, S. Botanical Pesticides: Use of Plants in Pest Management. CGC International Journal of Contemporary Technology and Research. 2021, 4(1), 271-275. |
| [34] | Pauly A., Breat, Y., Tchibozo, S., Aikpe, C. & Boeve, J. L. Hymenoptera Apoidea et Braconidae de quelques forêts sacrées du Sud-Bénin. Bulletin de la Société Royale Belge d'Entomologie/Bulletin van de Koninklijke Belgische Vereniging voor Entomologie. 2009, 45, 121-129. |
| [35] | Kwapong, P. K., Danquah, P. O. A., Asare, A. T. Insect floral visitors of cowpea (Vigna unguiculata L.). Annals of Biological Research. 2013, 4, 12-18. |
| [36] | Ige, O. E., Olotuah, O. F., Akerele, V. Floral biology and pollination ecology of cowpea (Vigna unguiculata L. Walp). Modern Applied Science. 2011, 5(4): 74-82. |
| [37] | Mohammadou, M., Fouelifack-Nintidem, B., Adamou, M., Taimanga, Kossini, D., Tsekane, S. J., Ngamaleu-Siewe, B., Kenne, E. L., Yomon, A. K., Kenne, M.. Diversity and Abundance of Pest Insects Associated with Vigna unguiculata (L.) Walp., 1843 (Fabales: Fabaceae) in Bockle and Dang Localities (North-Cameroon). American Journal of Entomology. 2023b, 7(2), 38-61. |
| [38] | Kosini, D., Nukenine, E. N., Tofel, K. H., Goudoungou, J. W., Langsi, D. J., Adamou, M., Abdou, J. P., Djafsia, B., Ndouwe, H. M. T. Impact of environment on Callosobruchus maculatus (Coleoptera: Chrysomelidae) response to acetone extract of Gnidia kaussiana Meisn (Thymeleaceae) and Ocimum canum Sim (Lamiaceae) botanical insecticides. European Journal of Nutrition and Food Safety. 2020, 12(8), 128-139. |
| [39] |
Tunsi, G. Les différentes zones agroécologiques du Cameroun. Les Journées Nationales Géonumériques de l’AFIGEO & DECRIPTAGEO. Centre des Congrès, 13 & 14 Septembre 2023, Reims. Available from:
http://www.limko.cm/gis-geomatics/2021/06/les-zones-agro-ecologiques-du-cameroun/ (accessed on 13 August 2023). |
| [40] | Mope, J. Rapport annuel d’activités du Service Provincial du Développement Communautaire de l’Adamaoua. Service Provincial du Développement Communautaire de l’Adamaoua (éd.); Ngaoundéré; 1997, pp. 1-95. |
| [41] | Letouzey, R. Etude phytogéographique du Cameroun. 5ème édition. Paris: Paul Le Chevalier; 1968, pp. 1-511. |
| [42] | Djoufack-Meneta, V. Étude multiéchelle des précipitations et du couvert végétal au Cameroun: Analyses spatiales, tendances temporelles, facteurs climatiques et anthropiques de variabilité. Thèse de Doctorat d’État, Université de Yaoundé I - Université de Bourgogne, 2011. |
| [43] |
Climate-Data.org. Ngaoundere Climate (Cameroon). Available from:
https://en.climate-data.org/africa/cameroon/adamawa/ngaoundere-898011/ (accessed on 13 August 2023). |
| [44] | Kottek, M., Grieser, J., Beck, C., Rudolf, B., Rubel, F. World Map of the Köppen-Geiger Climate Classification Updated. Meteorologische Zeitschrift. 2006, 15(3), 259-263. |
| [45] | Yaouba, B., Bitondo, D. Analysis of rainfall dynamics in the three main cities of northern Cameroon. Research Square. 2022, 12(12), 1-15. |
| [46] | MINEF (Ministry of Environment and Forestry (Cameroon),. Diagnostic général de la situation de l’environnement dans la province de l’Adamaoua. Document de base; PNUD/GTZ/BM; 1994, pp. 1-143. |
| [47] |
Sehou, A. Esclavage, émancipation et citoyenneté dans les lamidats de l’Adamaoua (Nord-Cameroun). Slaveries & Post-slaveries, 2019. Available from:
http://journals.openedition.org/slaveries/580 (accessed on 20 April 2024). |
| [48] | Sreekanth. Field evaluation of certain leaf extracts for the control of mussel scale (Lepidosaphes piperis Gr.) in Black pepper (Piper nigrum L.). Journal of Biopesticides. 2013, 6(1), 1-5. |
| [49] | Tchuenguem Fohouo, F.-N. Foraging and pollination behaviour of Apis mellifera adansonii Latreille (Hymenoptera: Apidae: Apinae) on the flowers of three plants in Ngaoundere (Cameroon): Callistemon rigidus (Myrtaceae), Syzygium guineense var. macrocarpum (Myrtaceae) and Voacanga africana (Apocynaceae). Doctorate Thesis, University of Yaounde I, Cameroon, 2005. |
| [50] | Jean-Prost, 1987. L’Apiculture. 6ème édition, Paris: Lavoisier; 1987, pp. 1-579. |
| [51] | Jacob-Remacle, A. Comportement de butinage de l'abeille domestique et des abeilles sauvages dans des vergers de pommiers en Belgique. Apidologie. 1989, 20(4), 271-285. |
| [52] | Rice, W. Analyzing tables of statistical tests. Evolution, 1989, 43(1), 223-225. |
| [53] | Mokam, D. G., Djieto-Lordon, C., Bilong Bilong, C. F. Patterns of species richness and diversity of insects associated with cucurbit fruits in the southern part of Cameroon. Journal of Insect Science. 2014, 2014, 14, 248. |
| [54] | Feng, B., Qian, K., Du, Y.-J. Floral Volatiles from Vigna unguiculata. Are Olfactory and Gustatory Stimulants for Oviposition by the Bean Pod Borer Moth Maruca vitrata. Insects. 2017, 8(2), 60. |
| [55] | Basualdo M., Bedascarrasbure E. & De Jong D. Africanized honey bees (Hymenoptera: Apidae) have a greater fidelity to sunflowers than European honey bees. Journal of Economic Entomology. 2000, 93, 304-307. |
| [56] | Wright, G. A., Skinner B. D., Smith B. H. Ability of honeybee, Apis mellifera, to detect and discriminate odors of varieties of canola (Brassica rapa and Brassica napus) and snapdragon flowers (Antirrhinum majus). Journal of Chemical Ecology. 2002, 28, 721-740. |
| [57] | Rucker, R. R., Thurman, W. N., Burgett, M. Honey bee pollination markets and the internalization of reciprocal benefits. American Journal of Agricultural Economics. 2012, 94(4), 956-977. |
| [58] | Fuzaro, L., Xavier, N. L., Carvalho, F. J., Silva, F. A. N., Carvalho, S. M., Andaló, V. Influence of pollination on canola seed production in the Cerrado of Uberlândia, Minas Gerais State, Brazil. Acta Scientiarum. Agronomy. 2018, 40: e39315. |
| [59] | Kengni, B. S., Ngakou, A., Tchuenguem Fohouo, F.-N.. Pollination and yield attributes of (cowpea) Vigna unguiculata L. Walp. (Fabaceae) as influenced by the foraging activity of Xylocopa olivacea Fabricius (Hymenoptera: Apidae) and inoculation with Rhizobium in Ngaoundere, Cameroon. International Journal of Agronomy and Agricultural Research. 2015, 6(2), 62-76. |
| [60] | Mbogning, E., Tchoumboue, J., Damesse, F., Sanou, S. M., Antonella, C. Caractéristiques physico-chimiques des miels de la zone Soudano-guinéenne de l’Ouest et de l’Adamaoua Cameroun. Tropicultura. 2011, 29(3), 168-175. |
| [61] | Gruber, M., Sanda, M. Honey Hunting and Beekeeping in Adamaoua (Cameroon). Brückner D. (ed.). Topics in Interdisciplinary African Studies 51. Köln, Germany: Rüdiger Köppe Verlag,; 2019, pp. 1-108. |
| [62] | Nchoutnji, I., Fofiri Nzossie, E. J., Olina Bassalan J.-P., Templen, L., Kamenin, A. Systèmes maraîchers en milieux urbain et périurbain des zones Soudano-sahélienne et Soudano-guinéenne du Cameroun: cas de Garoua et Ngaoundéré. Tropicultura. 2009, 2009, 27, 2, 98-104. |
| [63] | Fouepe Takounjou, A., Foano Dandjio, W., Fodoue, Y., Fantong, W., Ngah, M., Jokam Nenkam, T., Carlier C., Vassolo, S., Montcoudiol, N., Enow Tarkang, C., Chounna, G., Kringel, R. Assessment of water availability for agricultural activities in the savannah Plateau of Adamawa-Cameroon. Environ Monit Access. 2022, 194(11), 813. |
| [64] | Anderson, N. L., Harmon-Threatt, A. N. Chronic contact with realistic soil concentrations of imidacloprid affects the mass, immature development speed, and adult longevity of solitary bees. Scientific Reports. 2019, 9, 3724. |
| [65] |
Baskaran, S., Kookana, R. S., Naidu, R. Degradation of bifenthrin, chlorpyrifos and imidacloprid in soil and bedding materials at termiticidal application rates. Pesticide Science. 1999, 55, 1222–1228.
https://doi.org/10.1002/(SICI)1096-9063(199912)55:12<1222::AID-PS83>3.0.CO;2-7 |
| [66] |
Gervais, J. A., Luukinen, B., Buhl, K., Stone, D. Imidacloprid General Fact Sheet. National Pesticide Information Center (NPIC), Oregon State University Extension Services, 2010. Available from:
http://npic.orst.edu/factsheets/imidagen.html (accessed on 19 April 2022). |
| [67] | Bonmatin, J. M., Moineau, I., Charvet, R., Colin, M. E., Fleche, C., Bengach, E. R. Behaviour of imidacloprid in fields. Toxicity for honey bees. In Environmental Chemistry. Lichtfouse, E., Schwarzbauer, J., Robert, D., eds., Springer, Berlin Heidelberg, 2005, pp. 483–494. |
| [68] | Sharma, S., Singh, B. Metabolism and persistence of imidacloprid in different types of soils under laboratory conditions. International Journal of Environmental Analytical Chemistry. 2014, 94, 1100-1112. |
| [69] |
Wagner, S. Environmental fate of imidacloprid. California Department of Pesticide Regulation (CDPR), 2016. Available from;
https://www.cdpr.ca.gov/docs/emon/pubs/envfate.htm (accessed on 21 April 2024). |
| [70] | Hill, B. D., Inaba, D. J. Dissipation of lambda-Cyhalothrin on Fallow vs Cropper Soil. Journal of Agricultural and Food Chemistry, 1991, 39, 2282-2284. |
| [71] | Hornsby, A. G., Wauchope, R. D., Herner, A. E. Pesticide Properties in the Environment. New York: Springer; 1995, pp. 1-132. |
| [72] |
National Pesticide Information Center. NPIC: Lambda-cyhalothrin (Technical Fact Sheet). Oregon State University. USA, 2001, pp. 1-6. Available from:
http://npic.orst.edu/ingred/aifact.html (accessed on 19 April 2022). |
| [73] | Fouelifack-Nintidem, B., Yetchom-Fondjo, J. A., Tsekane, S. J., Ngamaleu-Siewe, B., Kenne, E. L., Biawa Kagmegni, M., Tuekam-Kowa, P. S., Yomon, A. K., Kentsop-Tsafong, R. M., Dim-Mbianda, A. M., Kenne, M. Diversity and abundance of pest insects associated with the Ethiopian eggplant plants Solanum aethiopicum Linnaeus, 1756 (Solanaceae) in Balessing (West-Cameroon). American Journal of Entomology. 2021, 5(3), 70-91. |
| [74] | Louveaux, J. L’abeille domestique dans ses relations avec les plantes cultivées. In Pollinisation et productions végétales, Pesson, P., Louveaux J., eds., INRA, Paris; 1984, pp. 527-555. |
| [75] | Seeley, T. D., Camazine, S., Sneyd, J. Collective decision - making in honey bees: how colonies choose among nectar sources. Behavior Ecology Sociobiology. 1991, 28, 277-290. |
APA Style
Taïmanga, Mohammadou, M., Mbianda, P. A., Adamou, M., Youssoufa, O., et al. (2024). Impact of Plant Extracts on the Pollination Activity of Apis mellifera Linnaeus, 1758 (Hymenoptera: Apidae) on Flowers of Cowpea Variety Feekem, in Dang (Adamaoua, Cameroon). American Journal of Entomology, 8(2), 31-59. https://doi.org/10.11648/j.aje.20240802.12
ACS Style
Taïmanga; Mohammadou, M.; Mbianda, P. A.; Adamou, M.; Youssoufa, O., et al. Impact of Plant Extracts on the Pollination Activity of Apis mellifera Linnaeus, 1758 (Hymenoptera: Apidae) on Flowers of Cowpea Variety Feekem, in Dang (Adamaoua, Cameroon). Am. J. Entomol. 2024, 8(2), 31-59. doi: 10.11648/j.aje.20240802.12
AMA Style
Taïmanga, Mohammadou M, Mbianda PA, Adamou M, Youssoufa O, et al. Impact of Plant Extracts on the Pollination Activity of Apis mellifera Linnaeus, 1758 (Hymenoptera: Apidae) on Flowers of Cowpea Variety Feekem, in Dang (Adamaoua, Cameroon). Am J Entomol. 2024;8(2):31-59. doi: 10.11648/j.aje.20240802.12
@article{10.11648/j.aje.20240802.12,
author = {Taïmanga and Moukhtar Mohammadou and Pharaon Auguste Mbianda and Moïse Adamou and Ousmana Youssoufa and Boris Fouelifack-Nintidem and Andrea Sarah Kenne Toukem and Odette Massah Dabole and Oumarou Abdoul Aziz and Abraham Tchoubou-Sale and Abdel Kayoum Yomon and Sedrick Junior Tsekane and Martin Kenne},
title = {Impact of Plant Extracts on the Pollination Activity of Apis mellifera Linnaeus, 1758 (Hymenoptera: Apidae) on Flowers of Cowpea Variety Feekem, in Dang (Adamaoua, Cameroon)
},
journal = {American Journal of Entomology},
volume = {8},
number = {2},
pages = {31-59},
doi = {10.11648/j.aje.20240802.12},
url = {https://doi.org/10.11648/j.aje.20240802.12},
eprint = {https://article.sciencepublishinggroup.com/pdf/10.11648.j.aje.20240802.12},
abstract = {Synthetic pesticides present worldwide risks of contamination of humans, livestock and the environment due to the strong persistence and the toxic residues in fruits and vegetables. Natural biopesticides of local plant origin present low persistence and are the best alternative for the control of crop pests. In the Adamaoua region (Northern Cameroon), few studies exist concerning effects of botanical pesticides on the behavior of beneficial insects. Studies aimed to draw up a list of pollinating insects on flowers of Vigna unguiculata (L.) Walp., 1843 (Fabales: Fabaceae), in situations of treatment with botanical pesticides compared to the situation of the use of synthetic insecticide and to determine the effect of the biopesticides on the behavior of the main floricultural insects. Field investigations were carried out during two cowpea cultivation campaigns (June to September 2021 and June to October 2022) in Dang (suburb of Ngaoundere) on the effect of leaves extracts of local plant origin on the foraging behavior of Apis mellifera Linnaeus, 1758 (Hymenoptera: Apidae) and the main sap-sucking insect Aphis craccivora Koch, 1854 (Hemiptera: Aphididae). Forty-four cowpea plots of 4x3.5 m each distributed according to the randomized complete block model (four untreated plots as negative control, four plots treated with the synthetic insecticide Parastar (40EC 535/ 10/IN, 20 g/l of imidacloprid and 20 g/l of lamda-cyhalothrin) as positive control, and 36 experimental plots treated with three concentrations (10%, 20% and 30%) of aqueous leaves extract of Calotropis procera (Gentianales: Apocynaceae), Eucalyptus camaldulensis (Myrtales: Myrtaceae), and Tithonia diversifolia (Asterales: Asteraceae) respectively, made it possible to conduct four treatments: (1) flowers left to freely pollination, (2) flowers protected against pollinators, (3) flowers visited exclusively by Ap. mellifera and (4) flowers protected against insects. Among eight species (four orders, four families and seven genera) recorded on the flowers of V. unguiculata, the domestic bee Ap. mellifera was the most common and collected nectar and pollen. The control plots and those treated with 10% or 20% aqueous leaves extracts allowed the bee to carry out its activity. Plots treated with 30% extract of each plant and those treated with the synthetic insecticide Parastar, drastically altered the rhythm and speed of activity in Ap. mellifera foragers. This behavior became less coordinated and slow on treated plants. It would be wise to use 10% or 20% aqueous extracts as botanical insecticides and an alternative to the synthetic insecticide Parastar.
},
year = {2024}
}
TY - JOUR T1 - Impact of Plant Extracts on the Pollination Activity of Apis mellifera Linnaeus, 1758 (Hymenoptera: Apidae) on Flowers of Cowpea Variety Feekem, in Dang (Adamaoua, Cameroon) AU - Taïmanga AU - Moukhtar Mohammadou AU - Pharaon Auguste Mbianda AU - Moïse Adamou AU - Ousmana Youssoufa AU - Boris Fouelifack-Nintidem AU - Andrea Sarah Kenne Toukem AU - Odette Massah Dabole AU - Oumarou Abdoul Aziz AU - Abraham Tchoubou-Sale AU - Abdel Kayoum Yomon AU - Sedrick Junior Tsekane AU - Martin Kenne Y1 - 2024/05/24 PY - 2024 N1 - https://doi.org/10.11648/j.aje.20240802.12 DO - 10.11648/j.aje.20240802.12 T2 - American Journal of Entomology JF - American Journal of Entomology JO - American Journal of Entomology SP - 31 EP - 59 PB - Science Publishing Group SN - 2640-0537 UR - https://doi.org/10.11648/j.aje.20240802.12 AB - Synthetic pesticides present worldwide risks of contamination of humans, livestock and the environment due to the strong persistence and the toxic residues in fruits and vegetables. Natural biopesticides of local plant origin present low persistence and are the best alternative for the control of crop pests. In the Adamaoua region (Northern Cameroon), few studies exist concerning effects of botanical pesticides on the behavior of beneficial insects. Studies aimed to draw up a list of pollinating insects on flowers of Vigna unguiculata (L.) Walp., 1843 (Fabales: Fabaceae), in situations of treatment with botanical pesticides compared to the situation of the use of synthetic insecticide and to determine the effect of the biopesticides on the behavior of the main floricultural insects. Field investigations were carried out during two cowpea cultivation campaigns (June to September 2021 and June to October 2022) in Dang (suburb of Ngaoundere) on the effect of leaves extracts of local plant origin on the foraging behavior of Apis mellifera Linnaeus, 1758 (Hymenoptera: Apidae) and the main sap-sucking insect Aphis craccivora Koch, 1854 (Hemiptera: Aphididae). Forty-four cowpea plots of 4x3.5 m each distributed according to the randomized complete block model (four untreated plots as negative control, four plots treated with the synthetic insecticide Parastar (40EC 535/ 10/IN, 20 g/l of imidacloprid and 20 g/l of lamda-cyhalothrin) as positive control, and 36 experimental plots treated with three concentrations (10%, 20% and 30%) of aqueous leaves extract of Calotropis procera (Gentianales: Apocynaceae), Eucalyptus camaldulensis (Myrtales: Myrtaceae), and Tithonia diversifolia (Asterales: Asteraceae) respectively, made it possible to conduct four treatments: (1) flowers left to freely pollination, (2) flowers protected against pollinators, (3) flowers visited exclusively by Ap. mellifera and (4) flowers protected against insects. Among eight species (four orders, four families and seven genera) recorded on the flowers of V. unguiculata, the domestic bee Ap. mellifera was the most common and collected nectar and pollen. The control plots and those treated with 10% or 20% aqueous leaves extracts allowed the bee to carry out its activity. Plots treated with 30% extract of each plant and those treated with the synthetic insecticide Parastar, drastically altered the rhythm and speed of activity in Ap. mellifera foragers. This behavior became less coordinated and slow on treated plants. It would be wise to use 10% or 20% aqueous extracts as botanical insecticides and an alternative to the synthetic insecticide Parastar. VL - 8 IS - 2 ER -
Department of Agronomy, Institute of Fisheries and Aquatic Sciences, University of Douala, Douala, Cameroon
Research Fields: Botanical insecticides, Bees, Biological agriculture, Yield agricultural products, Coton (Gossypium hirsutum), Pollinization
Laboratory of Biology and Physiology of Animal Organisms, University of Douala, Douala, Cameroon
Research Fields: Botanical insecticides, Bees, Biological agriculture, Yield agricultural products, Coton (Gossypium hirsutum), Pollinization
Laboratory of Biology and Physiology of Plant Organisms, University of Douala, Douala, Cameroon
Research Fields: Applied entomology, Insects biology, biostatistics and pollinators, Apiculture, Insects-plants interactions, Animal Ethology, Animal Ecology
Laboratory of Applied Zoology, Faculty of Sciences, University of Ngaoundere, Ngaoundere, Cameroon; Faculty of Medicine and Biomedical Sciences of Garoua, University of Garoua, Garoua, Cameroon
Research Fields: Botanical insecticides, Bees, Biological agriculture, Yield agricultural products, Coton (Gossypium hirsutum), Pollinization
Laboratory of Biology and Physiology of Animal Organisms, University of Douala, Douala, Cameroon
Research Fields: Botanical insecticides, Bees, Biological agriculture, Yield agricultural products, Coton (Gossypium hirsutum), Pollinization
Laboratory of Biology and Physiology of Animal Organisms, University of Douala, Douala, Cameroon
Research Fields: Pest control, Entomology, Biology of animal populations, Insects ecology, Applied entomology, Animal ethology
Laboratory of Biology and Physiology of Animal Organisms, University of Douala, Douala, Cameroon
Laboratory of Biology and Physiology of Plant Organisms, University of Douala, Douala, Cameroon
Research Fields: Botanical insecticides, Bees, Biological agriculture, Yield agricultural products, Coton (Gossypium hirsutum), Pollinization
Laboratory of Biology and Physiology of Animal Organisms, University of Douala, Douala, Cameroon
Research Fields: Biopesticides, Insect pest control, Biological agriculture, Bees, Biological agriculture, Pollinization
Laboratory of Biology and Physiology of Plant Organisms, University of Douala, Douala, Cameroon
Research Fields: Botanical insecticides, Bees, Biological agriculture, Yield agricultural products, Coton (Gossypium hirsutum), Pollinization
Laboratory of Biology and Physiology of Animal Organisms, University of Douala, Douala, Cameroon
Research Fields: Pest control, Entomology, Biology of animal populations, Insects ecology, Applied entomology, Animal ethology
Laboratory of Biology and Physiology of Animal Organisms, University of Douala, Douala, Cameroon
Research Fields: Applied zoology, Quality of life and biostatistics, Wildlife Protection, Control of protected areas, Animal Ethology, Animal Ecology
Laboratory of Biology and Physiology of Animal Organisms, University of Douala, Douala, Cameroon
Research Fields: Biostatistics, Biology of the Animal Populations, Entomology, Myrmecology, Animal Ethology, Animal Ecology, sociobiology, Applied entomology, plant protection, Biological control, pest insects
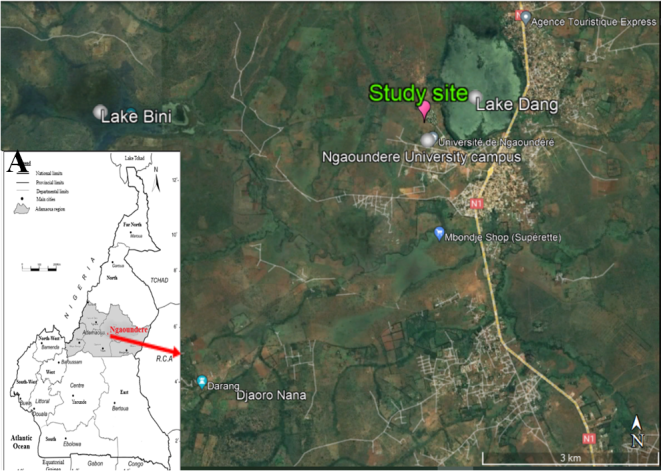
Figure 1. Localization map of the study site. A. Adamaoua Region in Cameroon (adapted from Sehou [47]); B. study site at Dang (Ngaoundere III suburb area) (Google Earth Pro for windows version 7.3.4.8642).

Figure 2. A: a cowpea plant 42 days after sowing; B: cowpea seeds or black-eyed bean “Feekem” variety from IRAD Garoua (North-Cameroon; C: a branch of Calotropis procera (Aiton) W. T. Aiton, 1811 (Gentianales: Apocynaceae) showing a blooming flower and green fleshy leaves; D: Eucalyptus camaldulensis Dehnh., 1832 (Myrtales: Myrtaceae); E: Some leaves and fruits of E. camaldulensis; F: a clump of Tithonia diversifolia (Hemsl.) A. Gray, 1883 (Asterales: Asteraceae) plants showing yellow blooming flowers.
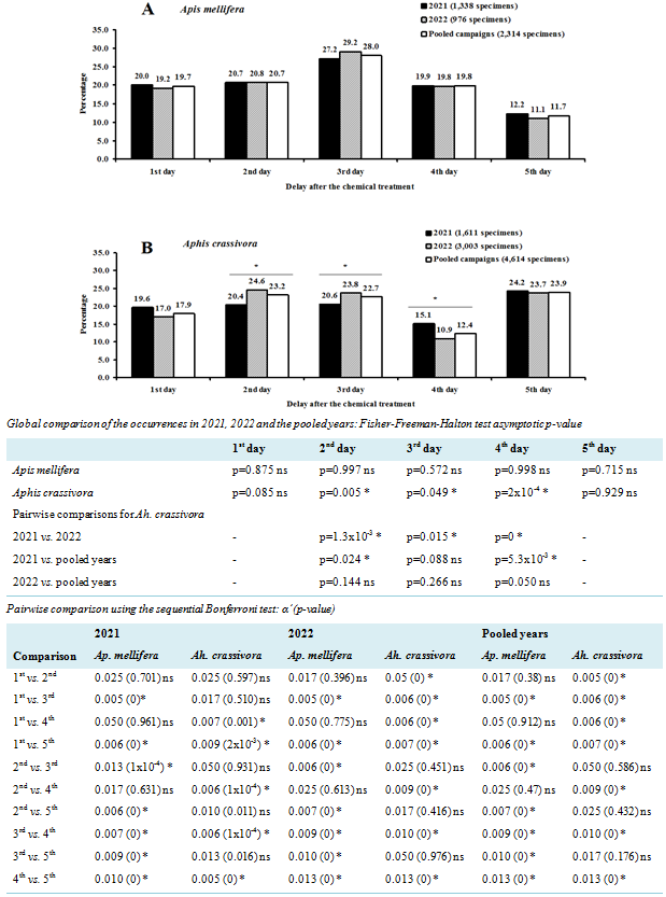
Figure 3. Evolution of the occurrence of Apis mellifera (Hymenoptera: Apidae) and Aphis crassivora (Hemiptera: Aphididae) on flowers of Vigna unguiculata (Fabales: Fabaceae) during five consecutive blooming days. ns: not significant difference (p>0.05); *: significant difference (p<0.05); α’: Bonferroni corrected significance level.
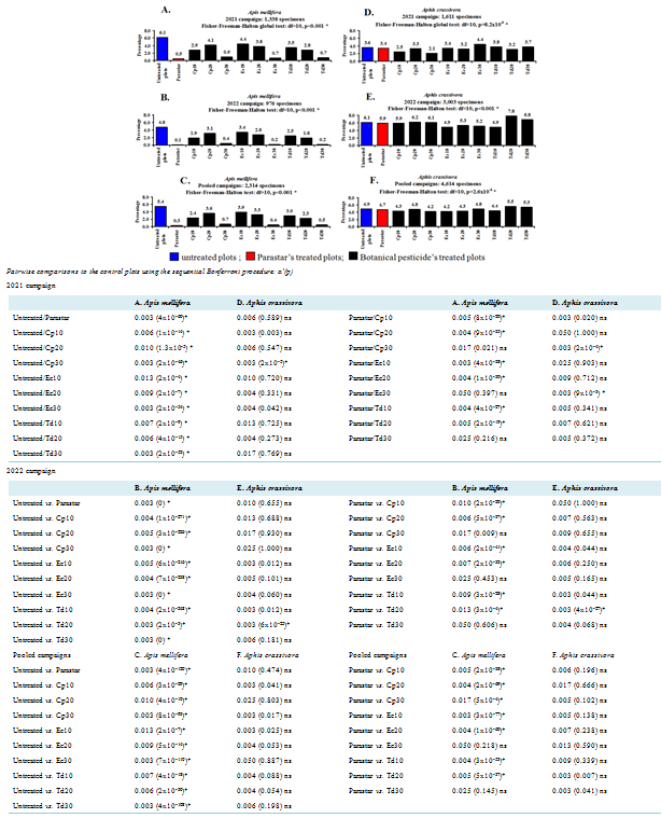
Figure 4. Percentages of Apis mellifera (Hymenoptera: Apidae) and Aphis crassivora (Hemiptera: Aphididae) on the flowers of Vigna unguiculata (Fabales: Fabaceae) in untreated plots or in plots treated using the synthetic insecticide (Parastar) and three aqueous leaves extracts of three plants. Cp10, Cp20 and Cp30: 10%, 20%, and 30%: concentrations of Calotropis procera (Gentianales: Apocynaceae); Ec10, EC20 and Ec30: 10%, 20%, and 30%: concentrations of Eucalyptus camaldulensis (Myrtales: Myrtaceae); Td10, Td20 and Td30: 10%, 20%, 30%: concentrations of Tithonia diversifolia (Asterales: Asteraceae). ns: not significant difference (p>α’); *: significant difference (p<0.05 or p<α’).

Figure 5. Apis mellifera (Hymenoptera: Apidae) worker harvesting pollen (A) and nectar (B) from a Vigna unguiculata flower in Dang.
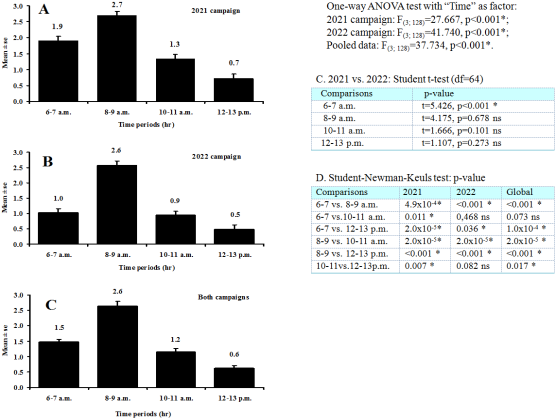
Figure 6. Daily rhythm of activity of Apis mellifera (Hymenoptera: Apidae) (33 trips each time period, 132 essays each campaign, 264 essays for the pooled data) in 2021 and 2022 campaigns in Dang.
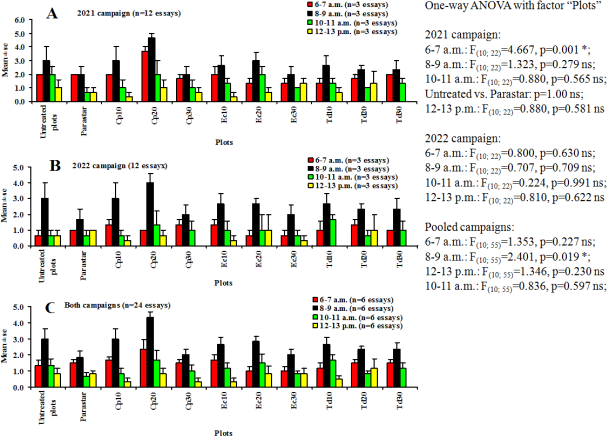
Figure 7. Rhythm of activity of Apis mellifera (Hymenoptera: Apida) on flowers of Vigna unguiculata (Fabales: Fabaceae). Cp10, Cp20 and Cp30: 10%, 20%, and 30%: concentrations of Calotropis procera; Ec10, EC20 and Ec30: 10%, 20%, and 30%: concentrations of Eucalyptus camaldulensis; Td10, Td20 and Td30: 10%, 20%, 30%: concentrations of Tithonia diversifolia. ns: not significant difference (p>α’); *: significant difference (p<0.05).
Information

