The Guinean Savanah Highlands of Adamawa is replete with multipurpose tree species, among which Burkea africana and Uvaria chamae are particularly noteworthy. Despite their importance, they remain in the wild and are subjected to overexploitation. The present study aims to contribute to the domestication of these species by root segment propagation. Specifically, the study aims to evaluate the seasonal variations in carbohydrate reserves (starch, soluble sugars, and sucrose) in other to determine the most favorable period for root cutting collection, assess the effect of substrate and length of root segment cuttings on the budding and rooting capacity of these species, evaluate the effect of carbohydrate content on bud emergence, root formation, callus induction, and control response in cuttings. For the seasonal fluctuation of carbohydrate, the experimental design was a complete randomized design with one factor represented by the season, and two replications. In the case the root propagation, the experimental design was a split-plot with three replications. The main treatment comprised three substrates (sand/sawdust, black soil/sawdust, black soil), while the sub-treatments were represented by three lengths of root segments cuttings (RSC) (10, 15, 20cm). The experimental unit consisted of 10 cuttings. Results showed that the onset of the rainy season coincides with peak of starch, soluble sugars and sucrose for Burkea africana and Uvaria chamae, marking the most favorable period for root cutting collection. The budding rate of Burkea africana showed significant variation (0.04 < 0.05), with values of 2.22 ± 1.96% in a black soil/sawdust mixture and 14.44 ± 12.36% in black soil after 26 weeks. For Uvaria chamae the best substrate was the mixture of black soil/sawdust (63.33 ± 45.27%). The number of leaves for Burkea africana was substantially higher in black soil (4.27 ± 2.16). The difference was statistically significant (0.008 < 0.01). For Burkea africana and Uvaria chamae, the optimal cutting length for budding was 20 cm (11.11 ± 10.52%, 81.11 ± 26.20%). The rooting rate of Uvaria chamae exhibited considerable variability the best rate was those of the black soil/sawdust substrate (22.22 ± 20.33%). The rooting rate varied from 5.55 ± 4.26% for cuttings of 10 cm to 23.33 ± 21.79% for those of 20 cm.Budded cuttings clustered with soluble sugars and sucrose. Rooted cuttings correlate negatively with all carbohydrate. Control cuttings were closely associated with starch. All these informations are important to develop scales and strategies toward the domestication of this species.
| Published in | American Journal of Agriculture and Forestry (Volume 14, Issue 2) |
| DOI | 10.11648/j.ajaf.20261402.11 |
| Page(s) | 74-91 |
| Creative Commons |
This is an Open Access article, distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution and reproduction in any medium or format, provided the original work is properly cited. |
| Copyright |
Copyright © The Author(s), 2026. Published by Science Publishing Group |
Burkea Africana, Uvaria Chamae, Guinean Savannah Highlands, Vegetative Propagation, RSC
Seasons | Dry season | Rainy season | P-value | ||
|---|---|---|---|---|---|
Beginning | End | Beginning | End | ||
Starch (g/100 g DM) | 5.65 ± 0.98bc | 4.63±0.59ab | 6.39±0.84c | 4.33±0.94a | 0.001 |
Seasons | Dry season | Rainy season | P-value | ||
|---|---|---|---|---|---|
Beginning | End | Beginning | End | ||
Starch (g/100 g DM) | 4.90 ± 0.64a | 7.32±0.87b | 8.49±0.73c | 4.57±0.49a | 0.0000 |
Seasons | Dry season | Rainy season | P-value | ||
|---|---|---|---|---|---|
Beginning | End | Beginning | End | ||
Soluble sugars (g/100 g DM) | 2.55 ± 0.31ab | 2.97±0.51c | 4.16±0.91d | 2.23±0.23a | 0.0000 |
Seasons | Dry season | Rainy season | P-value | ||
|---|---|---|---|---|---|
Beginning | End | Beginning | End | ||
Soluble sugars (g/100 g DM) | 3.72 ± 0.41b | 5.62±0.76c | 6.41±0.60d | 2.75±0.21a | 0.0000 |
Seasons | Dry season | Rainy season | P-value | ||
|---|---|---|---|---|---|
Beginning | End | Beginning | End | ||
Sucrose (g/100 g DM) | 1.34 ± 0.12a | 1.83±0.16b | 2.12±0.48b | 1.07±0.20a | 0.0000 |
Seasons | Dry season | Rainy season | P-value | ||
|---|---|---|---|---|---|
Beginning | End | Beginning | End | ||
Sucrose (g/100 g DM) | 2.78 ± 0.05b | 3.33±0.78c | 4.48±1.06d | 1.50±0.27a | 0.0000 |
Substrate | Number of shoots | Height of shoots | Number of leaves |
|---|---|---|---|
Sand/Sawdust | 1.46 ± 1.40b | 1.93 ± 1.66a | 2.27 ± 2.27b |
Black soil | 2.29 ± 1.43b | 2.49 ± 1.25a | 4.27 ± 2.16c |
Black soil/Sawdust | 0.52 ± 0.40a | 1.35 ± 1.10a | 0.91 ± 0.58a |
P-value | 0.02 | 0.59 | 0.007 |
Substrate | Number of shoots | Height of shoots | Number of leaves |
|---|---|---|---|
Sand/Sawdust | 23.44 ± 19.05a | 7.23 ± 6.12a | 76.11 ± 61.64a |
Black soil | 17.66 ± 17.67a | 5.63 ± 5.45a | 70.33 ± 69.97a |
Black soil/Sawdust | 18.88 ± 16.98a | 8.29 ± 5.68a | 92.11 ± 89.45a |
P-value | 0.31 | 0.48 | 0.81 |
Length of RSC (cm) | Number of shoots | Height of shoots (cm) | Number of leaves |
|---|---|---|---|
10 | 0.81 ± 0.46a | 0.65 ± 0.27a | 1.33 ± 1.08a |
15 | 1.81 ± 1.13a | 3.24 ± 2.84a | 3.25 ± 2.07a |
20 | 1.66 ± 1.52a | 1.88 ± 1.37a | 2.86 ± 2.59a |
P-value | 0.72 | 0.70 | 0.69 |
Length of RSC (cm) | Number of shoots | Height of shoots(cm) | Number of leaves |
|---|---|---|---|
10 | 8.33 ± 6.46a | 4.03 ± 2.27a | 26.22 ± 16.58a |
15 | 20.22 ± 15.57ab | 7.12 ± 5.85a | 91 ± 72.48b |
20 | 31.44 ± 17.57b | 10.00 ± 4.72a | 121.33 ± 78.68c |
P-value | 0.02 | 0.12 | 0.03 |
Substrates | Sand/Sawdust | Black soil | Black soil/Sawdust |
|---|---|---|---|
Mean | 14.44 ± 14.24a | 11.11 ± 10.64a | 22.22 ± 20.33a |
P-value | 0.44 | ||
Length of RSC (cm) | 10 | 15 | 20 |
|---|---|---|---|
Mean | 5.55 ± 4.26a | 18.88 ± 16.04a | 23.33 ± 21.79a |
P-value | 0.13 | ||
Substrates | Number of roots | Length of roots |
|---|---|---|
Sand/Sawdust | 2.88 ± 2.75a | 5.56 ± 3.07a |
Black soil | 1.55 ± 0.9a | 1.74 ± 1.22a |
Blacksoil/Sawdust | 2.77 ± 2.38a | 3.03 ± 2.54a |
P-value | 0.50 | 0.43 |
Lenght of RSC (cm) | Number of roots | Length of roots (cm) |
|---|---|---|
10 | 1 ± 0.22a | 1.67 ± 1.15a |
15 | 3.33 ± 2.5a | 5.31 ± 3.09a |
20 | 2.88 ± 2.42a | 3.33 ± 2.72a |
P-value | 0.16 | 0.12 |
Variables | Starch | Soluble sugars | Sucrose | Budding rate | Rooting rate |
|---|---|---|---|---|---|
Starch | 1 | ||||
Soluble sugars | 0,662 | 1 | |||
Sucrose | 0,864 | 0,949 | 1 | ||
Budding rate | -0,751 | -0,002 | -0,317 | 1 | |
Rooting rate | -0,947 | -0,867 | -0,980 | 0,500 | 1 |
Variables | Starch | Soluble sugars | Sucrose | Budding rate |
|---|---|---|---|---|
Starch | 1 | |||
Soluble sugars | -1,000 | 1 | ||
Sucrose | 1,000 | -1,000 | 1 | |
Budding rate | -1,000 | 1,000 | -1,000 | 1 |
RSC | Root Segment Cuttings |
| [1] | Neya, B., Hakkou, M., Pétrissans, M., & Gérardin, P., (2004). On the durability of Burkea africana heartwood: evidence of biocidal and hydrophobic properties responsible for durability. Annals of Forest Science, 61(3): 277-282 |
| [2] | Mulofwa, J., Simute, S., & Tengnäs, B., (1994). Agroforestry: manual for extension workers in Southern Province, Zambia. |
| [3] | Ezenyi, I. C., Okpoko, C. K., Ufondu, C. A., Okhale, S. E., & Adzu, B. (2021). Antiplasmodial, antinociceptive and antipyretic potential of the stem bark extract of Burkea africana and identification of its antiplasmodial-active fraction. Journal of traditional and complementary medicine, 11(4): 311-317. |
| [4] | Omulen L., Acere-lervick K., Ndyabarema R., Tumwijukyea, Asio S., (1997). District environment profile: Bushenyi. NORPLAN and Bushenyi District. Internal Report, 100 p. |
| [5] | Meunier Q., Bellefontaine R., Monteuuis O., (2008). Vegetative propagation of medicinal trees and shrubs for the benefit of rural communities in Uganda. Bois et Forêts des Tropiques, 295(2): 71-82. |
| [6] | Moupela, C., Doucet, J. L., Daïnou, K., Meunier, Q., & Vermeulen, C. (2013). Propagation trials by seed and air layering of Coula edulis Baill., and prospects for its domestication. Bois et Forêts des Tropiques., 318(4), 3-13 |
| [7] | Bellefontaine R., Meunier Q., Ichaou A., Morin A., Mapongmetsem P.M., Belem B., Azihou F., Houngnon A., Abdourhamane H., 2018. Regeneration by seeds and low-cost vegetative propagation (suckers and root segment cuttings) CIRAD. 460 p. |
| [8] | Ouédraogo E., Mando A., & Brussaard L., 2004. Soil macrofaunal-mediated organic resource disappearance in semi-arid West Africa. Applied Soil Ecology, 27(3): 259-267. |
| [9] | Bellefontaine R., Monteuuis O., (2002). Le drageonnage des arbres hors forêt: un moyen pour revégétaliser partiellement les zones arides et semi-arides sahéliennes ? In Verger M. (Ed) Multiplication végétative des ligneux forestiers, fruitiers et ornementaux. Montpellier, France: Cirad-Inra. Cirad |
| [10] | Hannah J., Jan Beniest., (2003). La multiplication végétative des ligneux en agroforesterie. Manuel de formation et bibliographie,162 p. |
| [11] | Mapongmetsem P.M., Njomba E., Fawa G., Oumarou Z., Dangai Y., Bellefontaine R., (2017). Vegetative Propagation of Vitex doniana Sweet from Root Segments Cuttings: Effects of Substrate and Length of Cuttings on the Rooting Ability. Annals of Experimental Biology, 5(1): 18-24. |
| [12] | Letouzey R., (1968). Phytogéographie du Cameroun. Edition Lechevalier, 518p. |
| [13] | Ministère de l’Environnement et des Forêts (MINEF), (1994). Diagnosis of the environment. Rio World Summit;.p. 113. |
| [14] | Yonkeu S., 1983. Végétation des pâturages de l’Adamaoua (Cameroun): écologie et potentialités pastorales. Thèse de Doctorat. Univ. Rennes I, France. 207p. |
| [15] | Mapongmetsem P.M., Alium P.S., Raouguedam J., Bawa K.L., Fawa G., (2016a). Vegetative propagation of Sclerocarya birrea, (A. Rich.) Hochst. from root segments cuttings: effect of substrate and root diameter. Annals of Experimental Biology, 4(2): 23-32. |
| [16] | Miller, G. L. (1972). Use of dinitrosalicylic acid reagent for determination of reducing sugars. Analytical Chemistry, 31, 426–428. |
| [17] | Dubois, M., Gilles, K. A., Hamilton, J. K., Rebers, P. A., & Smith, F. (1956). Colorimetric method for determination of sugars and related substances. Analytical Chemistry, 28(3), 350–356. |
| [18] | Jarvis, C. E., & Walker, J. R. (1993). Simultaneous, rapid, spectrophotometric determination of total starch, amylose and amylopectin. Journal of the Science of Food and Agriculture, 63(1): 53-57. |
| [19] | Ky-Dembélé C., Tigabu M., Bayala J., Savadogo P., Boussim I.J., Oden P.C., (2010). Clonal propagation of Detarium microcarpum from root cuttings. Silva Fennica, 44(5): 775-787. |
| [20] | Mapongmetsem P. M., Fawa G., Noubissie-Tchiagam J. B., Nkongmeneck B. A., Biaou S. S. H., Bellefontaine R., (2016 b). Vegetative propagation of Vitex doniana Sweet from root segments cuttings. Bois et Forêts des Tropiques, 327(1): 29 – 37. |
| [21] | Oumarou Z.H., Hamaya Y., Tsobou R., Abdoulaye H., Bellefontaine R., Mapongmetsem P.M., (2018). Vegetative propagation of Securidaca longepedunculata Fresen by cutting root segments. Afrique Science, 14(6): 388-399. |
| [22] | Stenvall, N., Piisilä, M., & Pulkkinen, P. (2009). Seasonal fluctuation of root carbohydrates in hybrid aspen clones and its relationship to the sprouting efficiency of root cuttings. Canadian Journal of Forest Research, 39(8), 1531-1537. |
| [23] | Sami, F., Yusuf, M., Faizan, M., Faraz, A., & Hayat, S., 2016. Role of sugars under abiotic stress. Plant physiology and biochemistry, 109, 54-61. |
| [24] | Coates palgrave, K (2002). Tree of southern Africa.Struik, Cape town,1212 pp. |
| [25] | Frost, P. (1996). The ecology of miombo woodlands. The miombo in transition: Woodlands and welfare in Africa, 266, 11-57. |
| [26] | Burkill, H. M. (1985). The Useful Plants of West Tropical Africa. Volume 1, Families A–D (2nd edition). Royal Botanic Gardens, Kew. Vol. 1: 960 p. |
| [27] | Landhäusser, S. M., & Lieffers, V. J., (2003). Seasonal changes in carbohydrate reserves in mature northern Populus tremuloides clones. Trees, 17(6): 471-476. |
| [28] | Oumarou H. Z., Hamawa Yougouda, Tsobou R., Dangai Yohana, Binwe J. B., Madi Amedi Damba R., Abdoulaye Herbert, Wangbitching J. D. D., Fawa Guidawa, Mapongmetsem P. M., (2019). Vegetative Propagation of Bombax costatum Pellegr. & Vuillet (Malvaceae) by Root Segments Cuttings: Effects of Mother Tree Diameter and Origin of Cuttings. American Journal of Agriculture and Forestry, 7(6): 248-258. |
| [29] | Zida, D., Tigabu, M., Sawadogo, L., Tiveau, D., & Odén, P. C. (2009). Long‐term effects of prescribed early fire, grazing and selective tree cutting on seedling populations in the Sudanian savanna of Burkina Faso. African Journal of Ecology, 47(1), 97-108. |
| [30] | Krishnamurthy, Kulithalai & Bahadur, Bir & Adams, Sebastian & Venkatasubramanian, Padma. (2015). Meristems and Their Role in Primary and Secondary Organization of the Plant Body. 151p. |
| [31] | Vanneste, S., Pei, Y., & Friml, J. (2025). Mechanisms of auxin action in plant growth and development. Nature Reviews Molecular Cell Biology, 26(10): 648–666. |
| [32] | Ky-Dembele, Catherine & Traoré, Fatoumata & B., Koné & Bayala, Jules & Antoine, Kalinganire & Bonneville, Jean & Olivier, Alain. (2015). Le bouturage est-il une option envisageable au Sahel ? Sahel Agroforesterie. 20. 4-5. |
| [33] | Belem B., Boussim J.I., Bellefontaine R., Guinko S., (2008). Stimulation du drageonnage de Bombax costatum Pelegr. et Vuillet par blessures de racines au Burkina Faso. Bois et Forêts des Tropiques, 295(1): 71-79. |
| [34] | Binwe J.B., Hamawa Y., Wangbitching J.D.D., Madi A.D.R., Apana E.J.H., Oumarou H.Z., Abdoulaye H., Fawa G., Mapongmetsem P. M., (2024). Influence of substrate and length on the ability of root segments cuttings of Ximenia americana L. to regenerate. International Journal of Research in Agronomy 7(9): 106-113. |
| [35] | Wangbitching J. D D, Yougouda, H., Guidawa, F., Binwe, J. B., Madi Ameti Damba R., Apana Ewodo H. J., Wamba Sopgou D.P., Oumarou H. Z., Abdoulaye H., & Mapongmetsem P. M. P., 2024. Influence of Substrate and Length on The Ability of Root Segments Cuttings of Amblygonocarpus andongensis (Welw. ex Oliv.) Exell & Torre to Regenerate. Discoveries in Agriculture and Food Sciences, 12(6): 56-71. |
| [36] | Rowe, J.H., Topping, J.F., Liu, J. and Lindsey, K., 2016, Abscisic acid regulates root growth under osmotic stress conditions via an interacting hormonal network with cytokinin, ethylene and auxin. New Phytologist, 211: 225-239. |
| [37] | Houehounha R., Avohou H.T., Sinsin B., Tandjiekpon A.M., (2009). Approches de régénération artificielle de Daniellia oliveri (Rolfe) Hutchison et Dalziel. International Journal of Biological and Chemical Sciences, 3(1): 7-19 |
| [38] | Hartmann H.T., Kaster D.E., Davies F.T., Geneve R.L., (2004). Plant Propagation: Principles and Practices. 6th ed. Prentice Hall of India Private Limited, New Delhi, India, p.770. |
| [39] | Harivel A., Bellefontaine R., Ousmane B., (2006). Aptitude à la multiplication végétative de huit espèces forestières d‟intérêt au Burkina Faso. Bois et Forêts des Tropiques, 288(2): 39-50. |
| [40] | Leakey, R.R., & Simons, A.J. (1997). The domestication and commercialization of indigenous trees in agroforestry for the alleviation of poverty. Agroforestry Systems, 38, 165-176. |
| [41] | Robinson J.C., Schwabe W.W., 1977. Studies on regeneration of Apple cultivars from root cuttings. Propagation aspects. Journal of Horticultural Science, 52(2): 205–220. |
| [42] | Stenvall N., Haapala T., PulkkinenP., (2004). Effect of genotype, age and treatment of stock plants on propagation of hybrid aspen (Populus tremula x Populus tremuloides) by root cuttings. Scandinavian Journal of Forest Research, 19(4): 303–311. |
| [43] | Magingo, F. S. S., & McP. Dick, J. (2001). Propagation of two miombo woodland trees by leafy stem cuttings obtained from seedlings. Agroforestry systems, 51(1), 49-55. |
| [44] | Fagge A. A. & Manga A. A., (2011). Effect of sowing media and gibberellic acid on the growth and seedling establishment of Bougainvillea glabra, Ixora coccinea and Rosa chinensis. 2. Root Characters. Bayero Journal of Pure and Applied Sciences, 4(2): 155-159. |
| [45] | Bisognin, D. A., Kielse, P., Lencina, K. H., & Mello, U. S. D. (2020). Vegetative propagation of Cordia trichotoma (Vell.) arrab. ex steud. by cuttings from shoots and roots. Cerne, 26, 265-271. |
| [46] | Da Costa, C. T., de Almeida, M. R., Ruedell, C. M., Schwambach, J., Maraschin, F. S., & Fett-Neto, A. G. (2013). When stress and development go hand in hand: Main hormonal controls of adventitious rooting in cuttings. Frontiers in Plant Science, 4, 133. |
| [47] | Thorpe, T. A. 2007. History of plant tissue culture. Molecular Biotechnology, 37(2): 169–180 |
| [48] | Eveland, A. L., & Jackson, D. P. (2012). Sugars, signalling, and plant development. Journal of experimental botany, 63(9), 3367-3377. |
| [49] | Wojtusik T., Boyd M. T. and Felker P., 1994: Effects of Different Media on Vegetative Propagation of Prosopic Cuttings Under Solar Power. Journal of Forest Ecology and Management, 69(1-3): 26-71. |
| [50] | Rosa M., Prado C., Podazza G., Interdonato R., González J.A., Hilal M., Prado F.E., (2009). Soluble sugars--metabolism, sensing and abiotic stress: a complex network in the life of plants. Plant Signal Behav; 4(5): 388-93 |
| [51] | Ciereszko, I. (2018). Regulatory roles of sugars in plant growth and development. Acta Societatis Botanicorum Poloniae, 87(2). |
| [52] | Bartolini G., Pestelli P., Toponi M. A. & Di monte G., 1996. Rooting and carbohydrate availability in Vitis 140 Ruggeri stem cuttings. Vitis, 35(1): 11–14. |
| [53] | MacNeill, G. J., Mehrpouyan, S., Minow, M. A., Patterson, J. A., Tetlow, I. J., & Emes, M. J. (2017). Starch as a source, starch as a sink: the bifunctional role of starch in carbon allocation. Journal of experimental botany, 68(16): 4433-4453. |
APA Style
Wangbitching, J. D. D., Dangai, Y., Oumarou, H. Z., Fawa, G., Binwe, J., et al. (2026). Interactive Effects of Substrate and Length on the Ability of Root Segment Cuttings of Burkea Africana (Hook) and Uvaria Chamae (P. Beauv) to Regenerate. American Journal of Agriculture and Forestry, 14(2), 74-91. https://doi.org/10.11648/j.ajaf.20261402.11
ACS Style
Wangbitching, J. D. D.; Dangai, Y.; Oumarou, H. Z.; Fawa, G.; Binwe, J., et al. Interactive Effects of Substrate and Length on the Ability of Root Segment Cuttings of Burkea Africana (Hook) and Uvaria Chamae (P. Beauv) to Regenerate. Am. J. Agric. For. 2026, 14(2), 74-91. doi: 10.11648/j.ajaf.20261402.11
AMA Style
Wangbitching JDD, Dangai Y, Oumarou HZ, Fawa G, Binwe J, et al. Interactive Effects of Substrate and Length on the Ability of Root Segment Cuttings of Burkea Africana (Hook) and Uvaria Chamae (P. Beauv) to Regenerate. Am J Agric For. 2026;14(2):74-91. doi: 10.11648/j.ajaf.20261402.11
@article{10.11648/j.ajaf.20261402.11,
author = {Jean de Dieu Wangbitching and Youhana Dangai and Haman Zepherin Oumarou and Guidawa Fawa and Jean-Baptiste Binwe and Rodrigue Damba Ameti Madi and Herve Joseph Ewodo Apana and Floriane Sorelle Eyenga and Pierre Marie Mapongmetsem},
title = {Interactive Effects of Substrate and Length on the Ability of Root Segment Cuttings of Burkea Africana (Hook) and Uvaria Chamae (P. Beauv) to Regenerate},
journal = {American Journal of Agriculture and Forestry},
volume = {14},
number = {2},
pages = {74-91},
doi = {10.11648/j.ajaf.20261402.11},
url = {https://doi.org/10.11648/j.ajaf.20261402.11},
eprint = {https://article.sciencepublishinggroup.com/pdf/10.11648.j.ajaf.20261402.11},
abstract = {The Guinean Savanah Highlands of Adamawa is replete with multipurpose tree species, among which Burkea africana and Uvaria chamae are particularly noteworthy. Despite their importance, they remain in the wild and are subjected to overexploitation. The present study aims to contribute to the domestication of these species by root segment propagation. Specifically, the study aims to evaluate the seasonal variations in carbohydrate reserves (starch, soluble sugars, and sucrose) in other to determine the most favorable period for root cutting collection, assess the effect of substrate and length of root segment cuttings on the budding and rooting capacity of these species, evaluate the effect of carbohydrate content on bud emergence, root formation, callus induction, and control response in cuttings. For the seasonal fluctuation of carbohydrate, the experimental design was a complete randomized design with one factor represented by the season, and two replications. In the case the root propagation, the experimental design was a split-plot with three replications. The main treatment comprised three substrates (sand/sawdust, black soil/sawdust, black soil), while the sub-treatments were represented by three lengths of root segments cuttings (RSC) (10, 15, 20cm). The experimental unit consisted of 10 cuttings. Results showed that the onset of the rainy season coincides with peak of starch, soluble sugars and sucrose for Burkea africana and Uvaria chamae, marking the most favorable period for root cutting collection. The budding rate of Burkea africana showed significant variation (0.04 Uvaria chamae the best substrate was the mixture of black soil/sawdust (63.33 ± 45.27%). The number of leaves for Burkea africana was substantially higher in black soil (4.27 ± 2.16). The difference was statistically significant (0.008 Burkea africana and Uvaria chamae, the optimal cutting length for budding was 20 cm (11.11 ± 10.52%, 81.11 ± 26.20%). The rooting rate of Uvaria chamae exhibited considerable variability the best rate was those of the black soil/sawdust substrate (22.22 ± 20.33%). The rooting rate varied from 5.55 ± 4.26% for cuttings of 10 cm to 23.33 ± 21.79% for those of 20 cm.Budded cuttings clustered with soluble sugars and sucrose. Rooted cuttings correlate negatively with all carbohydrate. Control cuttings were closely associated with starch. All these informations are important to develop scales and strategies toward the domestication of this species.},
year = {2026}
}
TY - JOUR T1 - Interactive Effects of Substrate and Length on the Ability of Root Segment Cuttings of Burkea Africana (Hook) and Uvaria Chamae (P. Beauv) to Regenerate AU - Jean de Dieu Wangbitching AU - Youhana Dangai AU - Haman Zepherin Oumarou AU - Guidawa Fawa AU - Jean-Baptiste Binwe AU - Rodrigue Damba Ameti Madi AU - Herve Joseph Ewodo Apana AU - Floriane Sorelle Eyenga AU - Pierre Marie Mapongmetsem Y1 - 2026/03/10 PY - 2026 N1 - https://doi.org/10.11648/j.ajaf.20261402.11 DO - 10.11648/j.ajaf.20261402.11 T2 - American Journal of Agriculture and Forestry JF - American Journal of Agriculture and Forestry JO - American Journal of Agriculture and Forestry SP - 74 EP - 91 PB - Science Publishing Group SN - 2330-8591 UR - https://doi.org/10.11648/j.ajaf.20261402.11 AB - The Guinean Savanah Highlands of Adamawa is replete with multipurpose tree species, among which Burkea africana and Uvaria chamae are particularly noteworthy. Despite their importance, they remain in the wild and are subjected to overexploitation. The present study aims to contribute to the domestication of these species by root segment propagation. Specifically, the study aims to evaluate the seasonal variations in carbohydrate reserves (starch, soluble sugars, and sucrose) in other to determine the most favorable period for root cutting collection, assess the effect of substrate and length of root segment cuttings on the budding and rooting capacity of these species, evaluate the effect of carbohydrate content on bud emergence, root formation, callus induction, and control response in cuttings. For the seasonal fluctuation of carbohydrate, the experimental design was a complete randomized design with one factor represented by the season, and two replications. In the case the root propagation, the experimental design was a split-plot with three replications. The main treatment comprised three substrates (sand/sawdust, black soil/sawdust, black soil), while the sub-treatments were represented by three lengths of root segments cuttings (RSC) (10, 15, 20cm). The experimental unit consisted of 10 cuttings. Results showed that the onset of the rainy season coincides with peak of starch, soluble sugars and sucrose for Burkea africana and Uvaria chamae, marking the most favorable period for root cutting collection. The budding rate of Burkea africana showed significant variation (0.04 Uvaria chamae the best substrate was the mixture of black soil/sawdust (63.33 ± 45.27%). The number of leaves for Burkea africana was substantially higher in black soil (4.27 ± 2.16). The difference was statistically significant (0.008 Burkea africana and Uvaria chamae, the optimal cutting length for budding was 20 cm (11.11 ± 10.52%, 81.11 ± 26.20%). The rooting rate of Uvaria chamae exhibited considerable variability the best rate was those of the black soil/sawdust substrate (22.22 ± 20.33%). The rooting rate varied from 5.55 ± 4.26% for cuttings of 10 cm to 23.33 ± 21.79% for those of 20 cm.Budded cuttings clustered with soluble sugars and sucrose. Rooted cuttings correlate negatively with all carbohydrate. Control cuttings were closely associated with starch. All these informations are important to develop scales and strategies toward the domestication of this species. VL - 14 IS - 2 ER -
Department of Biological Sciences, University of Ngaoundere, Ngaoundere, Cameroon
Higher Institute of Agriculture, Forestry, Water and Environment, University of Bertoua, Bertoua, Cameroon
Department of Plant Sciences, University of Bamenda, Bambili, Cameroon
Department of Science and Technique of Biological Agriculture, University of Ngaoundere, Ngaoundéré, Cameroon
Department of Biological Sciences, University of Ngaoundere, Ngaoundere, Cameroon
Department of Biological Sciences, University of Ngaoundere, Ngaoundere, Cameroon
Department of Biological Sciences, University of Ngaoundere, Ngaoundere, Cameroon
Department of Biological Sciences, University of Ngaoundere, Ngaoundere, Cameroon

Figure 1. Different length of cuttings.

Figure 2. Budded cuttings.
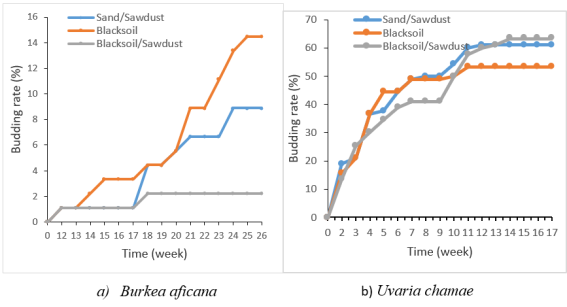
Figure 3. Effect of substrate on the budding rate.
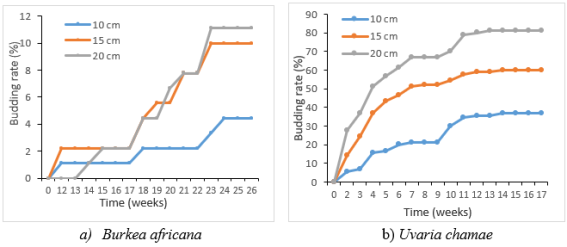
Figure 4. Effect of length of the cuttings on the budding rate.
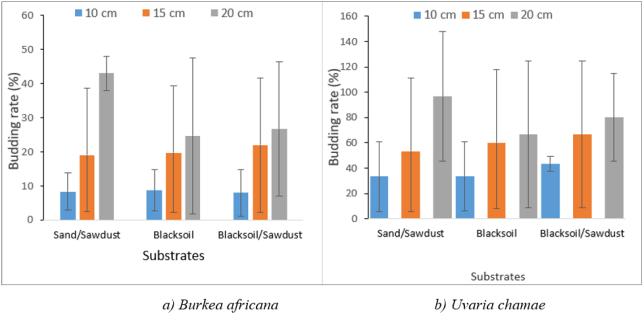
Figure 5. Budding rate following the interaction substrate*length of RSC.

Figure 6. Rooted cutting of Uvaria chamae.
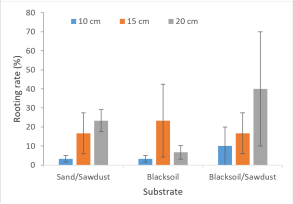
Figure 7. Rooting percentage according the interaction substrate*length.
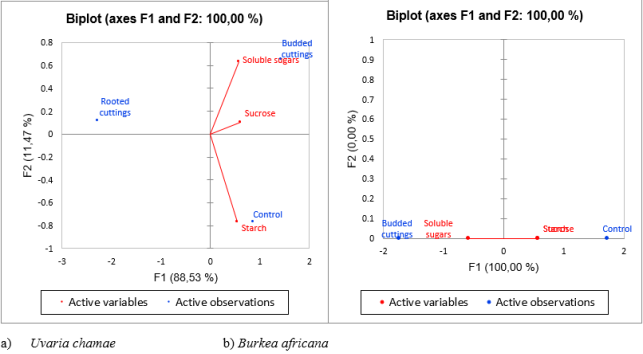
Figure 8. Principal Component Analysis (PCA) of Carbohydrate content and physiological state of different types of cuttings.
Information

