Ricinodendron heudelotii and Cola acuminata, are important plants species whose exploitation became abusive over the years due to the high utilization of their fruits as Non-Timber Forest Products (NTFP). They face to multiple challenges: a recalcitrance of seeds and salinity that limits regeneration. Therefore, regeneration seems an appropriate corridor for domestication with the optimization of plant mycorrhizal symbiosis otherwise called arbuscular mycorrhizal fungi (AMF). But alongside this domestication can be added constraints due to salinity of the soils in coastal region. This justify the aim of this work which was to study dynamics and evaluate the effect of salinity and mycorrhizal biofertilizers on the Ricinodendron heudelotii and Cola acuminata seedlings. To undergo this purpose, data were collected in two villages (Kendje and Njombeng) in Mungo division, and assay were conducted in greenhouse at the Faculty of Science, University of Douala-Cameroon. In the field, the identification of species was assessed over an area of 1600 m² as well as the circumference of the trees, the individual number of Ricinodendron heudelotii and Cola acuminata among other species in order to assess their maturity and rarity in the forest. The second part was carried out in the greenhouse for the purpose of germination, obtaining seedlings and evaluating the effects of arbuscular mycorrhizal fungi (Gisgaspora margarita) as biofertilizers on the seedlings in saline conditions (0, 50, 100 and 200 mM of NaCl). Some parameters were determined on seedlings (plant growth, dry weight, distribution of ions in plant organs, chlorophyll and carotenoid content) over a period of twenty-six weeks. Globally Cola acuminata is more present in the forest (5.88%) than Ricinodendron heudelotii (1.47%) with average circumference of 105cm for both species. AMF-biofertilizer alleviates the deleterious effect of salt stress on plants growth parameters depending of concentration. Moreover, for those species, the distribution of Na+ is more accumulated in the root’s plants unlike K+ and P which are more concentrated in the leaves.
| Published in | American Journal of Agriculture and Forestry (Volume 12, Issue 3) |
| DOI | 10.11648/j.ajaf.20241203.11 |
| Page(s) | 129-141 |
| Creative Commons |
This is an Open Access article, distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution and reproduction in any medium or format, provided the original work is properly cited. |
| Copyright |
Copyright © The Author(s), 2024. Published by Science Publishing Group |
Ecophysiology, Mycorrhizal Biofertilizer, Salinity, NTFP
2.1. Study Sites
2.2. Plant Material
2.3. Interaction Between Ricinodendron heudelotii, Cola acuminata and Their Environment
2.4. Trial Device
2.5. Fertilization
2.5.1. Plant Growth Parameters Determination
2.5.2. Assessment of Chlorophyll and Carotenoids Contents in Leaves
2.5.3. Biomass and Nutrients Determination
2.6. Statistical Analysis
3.1. Identification and Interaction of Plant Species in the Two Sites
3.2. Effects of Biofertilizer and Salt Stress on Growth Parameters of the Two Species
3.3. Effect of Biofertilizer and Salt Stress on Total Chlorophyll and Carotenoids Contents
3.4. Effect of NaCl and Biofertilizer on Some Physiological Parameters of Ricinodendron heudelotii and Cola acuminata
3.4.1. Nutrients Content in Different Parts of Ricinodendron heudelotii Seedlings
Amount (µg/gMS) | Organs | NaCl (mM) | ||||
|---|---|---|---|---|---|---|
0 | 50 | 100 | 200 | p | ||
N | Leaves | 1339.67 ± 51.67a | 3889.00±326.025c | 4196.00±0.00c | 3094.00±66.0b | p˂0.05 |
P | Leaves | 410.81±12.88a | 302.05±20.47a | 283.92±0.00a | 309.52±5.43a | p˃0.05 |
Stems | 233.41±7.32b | 161.50±10.94a | 166.49±.00a | 160.55±2.82a | p˃0.05 | |
Roots | 238.44±7.49b | 184.42±12.49a | 194.90±.00a | 190.83±3.35a | p˃0.05 | |
K+ | Leaves | 2052.15±63.93b | 1652.80±113.02a | 2627.48±0.00c | 2143.52±36.87b | p˂0.05 |
Stems | 1919.73±59.80b | 2091.83±143.04b | 1668.62±0.00a | 1816.58±31.24ab | p˃0.05 | |
Roots | 1855.23±57.79b | 1652.80±113.02b | 1549.91±0.00a | 1405.85±24.18a | p˃0.05 | |
Na+ | Leaves | 227.50 ± 8.75a | 197.75 ± 16.57a | 297.55 ±0.00b | 267.12 ± 5.68b | p˂0.05 |
Stems | 249.46 ± 9.60a | 238.38 ±19.99a | 249.47 ±0.00a | 244.27 ± 5.20a | p˃0.05 | |
Roots | 272.80± 10.49a | 309.29 ±25.93ab | 351.20 ±0.00b | 343.88 ± 7.31b | p˂0.05 | |
Amount (µg/gMS) | Organs | NaCl (mM) | ||||
|---|---|---|---|---|---|---|
0 | 50 | 100 | 200 | p | ||
N | Leaves | 1508.00±184.0b | 3972.00±.00b | 4279.67±164.83ab | 4139.67±170.60ab | p˂0.05 |
P | Leaves | 346.66±1413.3b | 324.00±.000b | 340.66±1311.45b | 259.6±1069.51a | p˃0.05 |
Stems | 610.00±5903.0c | 677.00±0.0b | 859.33±521.63a | 555.66±432.3c | p˂0.05 | |
Roots | 1015.6±2390.7a | 2408.0±2.92.3b | 1704.66±327.66b | 1478.0±721.0a | p˂0.05 | |
K+ | Leaves | 944.58±222.31b | 1757.60± 59.68c | 1513.87±29.11b | 703.54±73.34a | p˂0.05 |
Stems | 1022.02±240.5ab | 1297.93 ±44.07a | 1145.08 ± 22.02a | 1081.12±52.73b | p˃0.05 | |
Roots | 944.58±222.31a | 1519.58± 51.60b | 1145.08 ± 22.02a | 832.87±50.38a | p˃0.05 | |
Na+ | Leaves | 153.64±36.16b | 236.98 ± 9.47b | 231.97±4.46b | 186.80 ± 13.99a | p˂0.05 |
Stems | 153.64± 36.16b | 265.06 ± 9.00a | 278.15 ± 5.34bc | 312.80 ± 15.25c | p˂0.05 | |
Roots | 153.64 ± 36.16a | 336.77 ± 6.55c | 330.02 ± 6.34ab | 206.80 ± 13.99ab | p˂0.05 | |
3.4.2. Nutrients Content in Different Parts of Cola acuminata Seedlings
Amount (µg/gMS) | Organs | NaCl (mM) | ||||
|---|---|---|---|---|---|---|
0 | 50 | 100 | 200 | p | ||
N | Leaves | 2464.00±658.27a | 2149.00±622.56a | 1923.00±481.00a | 1848.67±288.702a | p˃0.05 |
P | Leaves | 117.48±34.03a | 155.18±24.24a | 273.22±72.98a | 141.78±35.44a | p˃0.05 |
Stems | 117.48±34.03a | 124.23±19.40ab | 218.98±58.50b | 134.20±33.55ab | p˃0.05 | |
Roots | 111.21±32.22a | 130.42±20.37a | 258.40 ± 69.03a | 142.79±35.70a | p˃0.05 | |
K+ | Leaves | 830.05±240.44a | 1298.0 ± 202.75a | 1940.76±518.44a | 1144.16±286.04a | p˃0.05 |
Stems | 359.02±104.00a | 1054.15±164.66ab | 1283.45±342.85b | 1017.84±254.46ab | p˃0.05 | |
Roots | 975.09±282.45a | 1054.15±164.66a | 1507.71±402.76a | 822.71±205.67a | p˃0.05 | |
Na+ | Leaves | 133.01±38.52a | 202.89±31.69a | 273.22±72.98a | 238.04±59.51a | p˃0.05 |
Stems | 108.81±31.52a | 244.57±38.20ab | 354.50±94.70b | 280.96±70.24ab | p˃0.05 | |
Roots | 193.23±55.97a | 244.57±38.20a | 354.50±94.70a | 238.04±59.51a | p˃0.05 | |
Amount (µg/gMS) | Organs | NaCl (mM) | ||||
|---|---|---|---|---|---|---|
0 | 50 | 100 | 200 | p | ||
N | Leaves | 1795 ± 422.54a | 2613.33 ± 88.62b | 2151.33 ± 41.33ab | 2350.67±114.66a | p˃0.05 |
P | Leaves | 138.13 ± 32.51a | 116.59 ± 11.43a | 169.75±3.26a | 106.93±9.60a | p˃0.05 |
Stems | 91.74 ± 21.59a | 138.66±4.70b | 167.82±3.22b | 155.00±7.59b | p˂0.05 | |
Roots | 101.57 ± 23.90a | 160.70±25.32b | 170.40 ± 3.27b | 147.82±7.21b | p˃0.05 | |
K+ | Leaves | 944.58±222.31a | 1757.60± 59.68b | 1513.87±29.11b | 1503.54±73.34b | p˂0.05 |
Stems | 1022.02±240.54a | 1297.93±44.07a | 1145.08±22.02a | 1081.12±52.73a | p˃0.05 | |
Roots | 944.58±222.31a | 1519.58±51.60a | 1145.08±22.02a | 1032.87±50.38a | p˃0.05 | |
Na+ | Leaves | 153.64 ± 36.16a | 236.98 ± 9.47b | 231.97±4.46b | 286.80±13.99b | p˂0.05 |
Stems | 153.64 ± 36.16a | 265.06 ± 9.00b | 278.15±5.34b | 212.80±15.25b | p˂0.05 | |
Roots | 153.64 ± 36.16a | 336.77 ± 6.55b | 330.02 ± 6.34b | 286.80 ± 13.99b | p˂0.05 | |
4.1. Identification of Species
4.2. Growth Parameters and Photosynthetic Pigments
4.3. Nutrients Content in Different Parts of Seedlings
| [1] | Ahammed G. J., Hajiboland R., 2024. Introduction to Arbuscular Mycorrhizal Fungi and Higher Plant Symbiosis: Characteristic Features, Functions, and Applications. In: Ahammed, G. J., Hajiboland, R. (eds) Arbuscular Mycorrhizal Fungi and Higher Plants. Springer, p: 1-17. |
| [2] | Atyi R. E., Devers D., De Wasseige C., Maisels F., 2009. « État des forêts d’Afrique centrale: synthèse sous-régionale ». Les forêts du Bassin du Congo- État des forêts 2008, p. 17-44, Luxembourg, Office des publications de l’Union européenne, 426 p ISBN 978-92-79-132 11-7. |
| [3] | Assogbadjo A E., Codjia T. C., Sinsin B., Ekue M. R., Mensoh G. A., 2005. Importance of rodents as a human food source in Benin, Belg. J. Zool. 135: 11-15. |
| [4] | Bago B., Pfeffer P. E., Shachar-Hill Y., 2000. Carbon metabolism and transport in arbuscular mycorrhizae. Plant Physiology 124(3): 949–958. |
| [5] | Barea N. J. M., Azcón R., Azcón-Aguilar G., 2002. Mycorrhizosphere interactions to improve plant fitness and soil quality. Anatonie van Leeuwenhoek, Springer Nature: 81(1–4): 343–351. |
| [6] | Benidire L., Daoui K., Fatemi Z. A., Achouak W., Bouarab L., OufdouK, 2015. Effect of salt stress on germination and seedling of Vicia faba L. Journal of Materials and Environmental Science. 6(3): 840-851. |
| [7] | Borde M., Dudhane M., Kulkarni M., 2017. Role of arbuscular mycorrhizal fungi (AMF) in salinity tolerance and growth response in plants under salt stress conditions. In Mycorrhiza-Eco-Physiology, Secondary Metabolites, Nanomaterials; Varma, A., Prasad, R., Tuteja, N., Eds.; Springer: Cham, Switzerland. 71–86. |
| [8] | CIFOR., 2010. Atelier sous-régional sur «l’Harmonisation des revues nationales sur les produits forestiers non ligneux (PFNL) en Afrique centrale», 17-18 Mai 2010, Douala, Cameroun, Rapport général: 26p. |
| [9] | Eke Balla M. S., 2021. The importance of social capital in the adoption of sustainable management practices of non-timber forest products (NTFPs) in Cameroon. African Scientific Journal, 3(4): 399-399. |
| [10] | FAO et PNUE, 2020. La situation des forêts du monde. Forêts, biodiversité et activité humaine. Rome, Italie: 11p. |
| [11] | Ferddermann N., Finlay R., Boller T., Elfstrand M., 2010. Functional diversity in arbuscular mycorrhizae. The role of gene expression phosphorous nutrition and symbiotic efficiency. Fungal Ecology 3(1): 1-8. |
| [12] | Giri, B., Kapoor R., Mukerji K. G., 2003. Influence of arbuscular mycorrhizal fungi and salinity on growth, biomass, and mineral nutrition of Acacia auriculiformis. Biology and Fertility of Soils 38: 170–175. |
| [13] | Haro H., Sanon K. B., Le Roux C., Duponnois R., Traoré A. S., 2020. Improvement of cowpea productivity by rhizobia and mycorrhizal inoculation in Burkina Faso. Symbiosis: 1-14. |
| [14] | Helgason T., Fitter A., 2005. The ecology and evolution of the Arbuscular Mycorrhizal Fungi. Mycologist 19(3): 96–101. |
| [15] | Henry E. E. Y., Kinsou E., Mensah A. C. G., Komlan F. A., Gandonou C. B., 2021. Réponse des plantes de tomate (Lycopersicon esculentum Mill.) cultivées sous stress salin à une application exogène de calcium et de potassium. Journal of Applied Biosciences 159(1): 16363-16370. |
| [16] | Kakabouki I., Stavropoulos P., Roussis I., Mavroeidis A., Bilalis D., 2023. Contribution of Arbuscular Mycorrhizal Fungi (AMF) in Improving the Growth and Yield Performances of Flax (Linum usitatissimum L.) to Salinity Stress. Agronomy 13, 2416. |
| [17] | Klinsukon C., Lumyong S., Kuyper T W., Boonlue S., 2021. Colonization by arbuscular mycorrhizal fungi improves salinity tolerance of eucalyptus (Eucalyptus camaldulensis) seedlings. Science Reports 11(1): 4362. |
| [18] | Malik J. A., Al-Qarawi A. A., Dar B. A., Hashem A., Alshahrani T. S., AlZain M. N., Habib., M. M., Javed M. M., Abd Allah E. F., 2022. Arbuscular Mycorrhizal Fungi Isolated from Highly Saline “Sabkha Habitat” Soil Alleviated the NaCl-Induced Stress and Improved Lasiurus scindicus Henr. Growth. Agriculture, 12(3): 337 |
| [19] | Ndonda A., Mahungu N., Moango A., Yandju M. C., 2019. Effet des champignons mycorhiziens Arbusculaires sur le phosphore des sols tropicaux et implication dans la biosynthèse du caroténoïde du manioc. J. Appl Biosciences 135: 13750-13765. |
| [20] | Ngonkeu M. E. L., 2003. Biodiversité et potentiel des mycorhizes à arbuscules de certaines zones agroécologiques du Cameroun. Thèse de Doctorat 3e cycle, Université de Yaoundé I. 258p. |
| [21] | Ngueuleu D. A., Muyang R. F., Wamba F. O., Ngotta B. B., Tefouet V. D., Nyemene K. P. J., Asseng C. C., Taffouo V. D., 2021. Evaluation of soils fertility, growth, nutrient uptake and yield traits of peanut under indigenous and effective microorganism fertilizers in sandy ferralitic soils in Douala, Cameroon. African Journal of Agricultural Research 17(3): 432-441. |
| [22] | Ngwasiri C., DjeR-Uam N., Vabi M., 2002. Legislative and institutional instruments for the sustainable management of non-timber forest product (NTFPs) in Cameroon. Past, present and unresolved issues. Projet de développement de foresterie communautaire, Yaoundé, Cameroun. Actualités des Forêts Tropicales OIBT, 32p. |
| [23] | Oumar C. S., Wade D., Faye E., Moustapha M. D., 2020. Effect of salt stress on the growth of four provenances of cashew tree (Anacardium occidentale L.) in a semi-controlled environment. VertigO - la revue électronique en sciences de l'environnement. |
| [24] | Ouaba J., Souaibou A., Niassy S., Lontsi-Demano M., Mayi M. P. A., Meutchieye F., Tchuinkam T., 2023. Socio-economic exploitation of edible caterpillars in the bimodal rainforest zone of Cameroon. Journal of Insects as Food and Feed: 1-12. |
| [25] | Shidiki A. A., Zanguim T G H., Tchamba N. M., 2021. Governance of Non-Timber Forest Products (NTFPs) Djansang (Ricinodendron heudelotii) and Wild Mango (Irvingia gabonensis) and Its Influence on the Livelihood of Rural Communities of Yokadouma in the East Region of Cameroon. Open Journal of Forestry, 11(2): 153-170. |
| [26] | Shapiro A., d’Annunzio R., Desclée B., Jungers Q., Kondjo H K., Iyanga J M., Kipute D D., 2023. Small scale agriculture continues to drive deforestation and degradation in fragmented forests in the Congo Basin 2015–2020. Land Use Policy, 134. |
| [27] | Sunderland, T. C. H., Harrison, S. T., Ndoye, O., 2004. Commercialisation of Non-timber Forest products in Africa: history, context and Prospects. in T. C. H. Sunderland and O. Ndoye (eds) Forest products, Livelihoods and conservation: Case-Studies of Non-Timber forest products Systems. Center for International Forestry Research Vol. 2: 23p. |
| [28] | Taibi K., Taibi F., Abderrahim L. A., Ennajah A., Belkhodja M., Mulet J. M., 2016. Effect of salt stress on growth, chlorophyll content, lipid peroxidation and antioxidant defence systems in Phaseolus vulgaris L. South African Journal of Botany 105: 306–312. |
| [29] | Talaat N B., Shawky B T., 2011. Influence of arbuscular mycorrhizae on yield, nutrients, organic solutes, and antioxidant enzymes of two wheat cultivars under salt stress. Journal of Plant Nutrition and Soil Science 174(2): 83–291. |
| [30] | Verhegghen A., Mayaux P., Wasseige C., Defourny P., 2012. Mapping Congo Basin vegetation types from 300 m and 1 km multi-sensor time series for carbon stocks and forest areas estimation. Biogeosciences, 9, 5061–5079. |
| [31] | Wacquant, J P., 1974. Recherche sur les propriétés d’absorption cationique des racines. Rôle Physiologique et importance écologique. Thèses de doctorat, Montpellier. 155p. |
| [32] | Xiao L., Duo Zheng., Naili Z., Lijia D., Aiping W., Qiqian W., Hua L., Mingshui Z., Yan L., Xinping W., Yanhong W., 2024. Arbuscular mycorrhizal fungi-mediated resistance to salt spray in Cinnamomum camphora seedlings enhanced by leaf functional traits. Soil Ecology Letters 6(3): |
| [33] | Youssef El-Iklil., Karrou M., Mrabet M., Benichou M., 2002. Effect of salt stress on the variation of certain metabolites in Lycopersicon esculentum and Lycopersicon sheesmanii. Canadian Journal of Plant Science 82(1): 177-183. |
APA Style
Virginie, T. I. A., Cyril, L. P. A., Serge, E. S., Ibrahim, F., Desire, T. V. (2024). Ecophysiology and Seedlings Nutrient Contents of Forest Species Ricinodendron heudelotii (Mull. Arg.) and Cola acuminata (P. Beauv.) Influenced by Biofertilizer and Salinity. American Journal of Agriculture and Forestry, 12(3), 129-141. https://doi.org/10.11648/j.ajaf.20241203.11
ACS Style
Virginie, T. I. A.; Cyril, L. P. A.; Serge, E. S.; Ibrahim, F.; Desire, T. V. Ecophysiology and Seedlings Nutrient Contents of Forest Species Ricinodendron heudelotii (Mull. Arg.) and Cola acuminata (P. Beauv.) Influenced by Biofertilizer and Salinity. Am. J. Agric. For. 2024, 12(3), 129-141. doi: 10.11648/j.ajaf.20241203.11
AMA Style
Virginie TIA, Cyril LPA, Serge ES, Ibrahim F, Desire TV. Ecophysiology and Seedlings Nutrient Contents of Forest Species Ricinodendron heudelotii (Mull. Arg.) and Cola acuminata (P. Beauv.) Influenced by Biofertilizer and Salinity. Am J Agric For. 2024;12(3):129-141. doi: 10.11648/j.ajaf.20241203.11
@article{10.11648/j.ajaf.20241203.11,
author = {Tchiaze Ifoue Alice Virginie and Lieujip Piengang Aurelien Cyril and Ekwel Sondi Serge and Fanteunti Ibrahim and Taffouo Victor Desire},
title = {Ecophysiology and Seedlings Nutrient Contents of Forest Species Ricinodendron heudelotii (Mull. Arg.) and Cola acuminata (P. Beauv.) Influenced by Biofertilizer and Salinity
},
journal = {American Journal of Agriculture and Forestry},
volume = {12},
number = {3},
pages = {129-141},
doi = {10.11648/j.ajaf.20241203.11},
url = {https://doi.org/10.11648/j.ajaf.20241203.11},
eprint = {https://article.sciencepublishinggroup.com/pdf/10.11648.j.ajaf.20241203.11},
abstract = {Ricinodendron heudelotii and Cola acuminata, are important plants species whose exploitation became abusive over the years due to the high utilization of their fruits as Non-Timber Forest Products (NTFP). They face to multiple challenges: a recalcitrance of seeds and salinity that limits regeneration. Therefore, regeneration seems an appropriate corridor for domestication with the optimization of plant mycorrhizal symbiosis otherwise called arbuscular mycorrhizal fungi (AMF). But alongside this domestication can be added constraints due to salinity of the soils in coastal region. This justify the aim of this work which was to study dynamics and evaluate the effect of salinity and mycorrhizal biofertilizers on the Ricinodendron heudelotii and Cola acuminata seedlings. To undergo this purpose, data were collected in two villages (Kendje and Njombeng) in Mungo division, and assay were conducted in greenhouse at the Faculty of Science, University of Douala-Cameroon. In the field, the identification of species was assessed over an area of 1600 m² as well as the circumference of the trees, the individual number of Ricinodendron heudelotii and Cola acuminata among other species in order to assess their maturity and rarity in the forest. The second part was carried out in the greenhouse for the purpose of germination, obtaining seedlings and evaluating the effects of arbuscular mycorrhizal fungi (Gisgaspora margarita) as biofertilizers on the seedlings in saline conditions (0, 50, 100 and 200 mM of NaCl). Some parameters were determined on seedlings (plant growth, dry weight, distribution of ions in plant organs, chlorophyll and carotenoid content) over a period of twenty-six weeks. Globally Cola acuminata is more present in the forest (5.88%) than Ricinodendron heudelotii (1.47%) with average circumference of 105cm for both species. AMF-biofertilizer alleviates the deleterious effect of salt stress on plants growth parameters depending of concentration. Moreover, for those species, the distribution of Na+ is more accumulated in the root’s plants unlike K+ and P which are more concentrated in the leaves.
},
year = {2024}
}
TY - JOUR T1 - Ecophysiology and Seedlings Nutrient Contents of Forest Species Ricinodendron heudelotii (Mull. Arg.) and Cola acuminata (P. Beauv.) Influenced by Biofertilizer and Salinity AU - Tchiaze Ifoue Alice Virginie AU - Lieujip Piengang Aurelien Cyril AU - Ekwel Sondi Serge AU - Fanteunti Ibrahim AU - Taffouo Victor Desire Y1 - 2024/05/10 PY - 2024 N1 - https://doi.org/10.11648/j.ajaf.20241203.11 DO - 10.11648/j.ajaf.20241203.11 T2 - American Journal of Agriculture and Forestry JF - American Journal of Agriculture and Forestry JO - American Journal of Agriculture and Forestry SP - 129 EP - 141 PB - Science Publishing Group SN - 2330-8591 UR - https://doi.org/10.11648/j.ajaf.20241203.11 AB - Ricinodendron heudelotii and Cola acuminata, are important plants species whose exploitation became abusive over the years due to the high utilization of their fruits as Non-Timber Forest Products (NTFP). They face to multiple challenges: a recalcitrance of seeds and salinity that limits regeneration. Therefore, regeneration seems an appropriate corridor for domestication with the optimization of plant mycorrhizal symbiosis otherwise called arbuscular mycorrhizal fungi (AMF). But alongside this domestication can be added constraints due to salinity of the soils in coastal region. This justify the aim of this work which was to study dynamics and evaluate the effect of salinity and mycorrhizal biofertilizers on the Ricinodendron heudelotii and Cola acuminata seedlings. To undergo this purpose, data were collected in two villages (Kendje and Njombeng) in Mungo division, and assay were conducted in greenhouse at the Faculty of Science, University of Douala-Cameroon. In the field, the identification of species was assessed over an area of 1600 m² as well as the circumference of the trees, the individual number of Ricinodendron heudelotii and Cola acuminata among other species in order to assess their maturity and rarity in the forest. The second part was carried out in the greenhouse for the purpose of germination, obtaining seedlings and evaluating the effects of arbuscular mycorrhizal fungi (Gisgaspora margarita) as biofertilizers on the seedlings in saline conditions (0, 50, 100 and 200 mM of NaCl). Some parameters were determined on seedlings (plant growth, dry weight, distribution of ions in plant organs, chlorophyll and carotenoid content) over a period of twenty-six weeks. Globally Cola acuminata is more present in the forest (5.88%) than Ricinodendron heudelotii (1.47%) with average circumference of 105cm for both species. AMF-biofertilizer alleviates the deleterious effect of salt stress on plants growth parameters depending of concentration. Moreover, for those species, the distribution of Na+ is more accumulated in the root’s plants unlike K+ and P which are more concentrated in the leaves. VL - 12 IS - 3 ER -
Department of Plant Biology, Faculty of Science, University of Douala, Douala, Cameroon
Department of Plant Biology, Faculty of Science, University of Douala, Douala, Cameroon
Department of Plant Biology, Faculty of Science, University of Douala, Douala, Cameroon
Department of Plant Biology, Faculty of Science, University of Douala, Douala, Cameroon
Department of Plant Biology, Faculty of Science, University of Douala, Douala, Cameroon

Figure 1. Pre-germination of Ricinodendron heudelotii (A) and Cola acuminata (B).
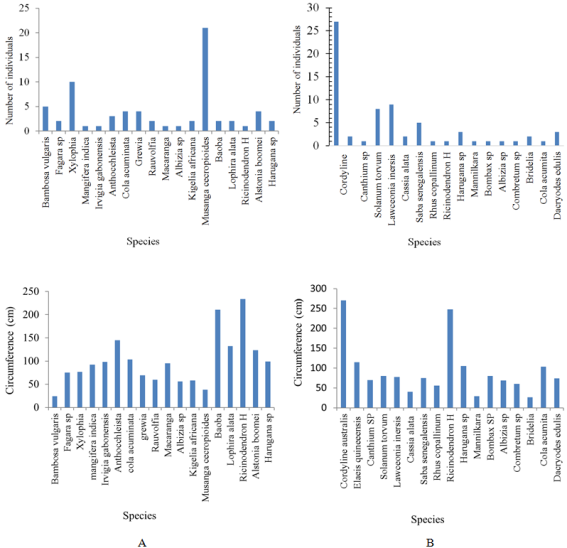
Figure 2. Number of individuals and their circumferences (A: Kendje; B: Njoumbeng).
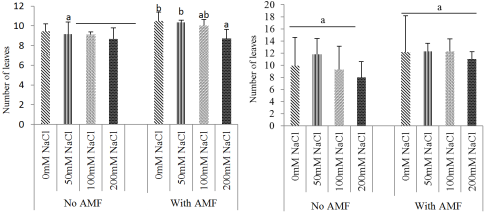
Figure 3. Effect of mycorrhizae inoculation on number of leaves: Ricinodendron heudelotii (A) and Cola acuminata (B) under salt stress.
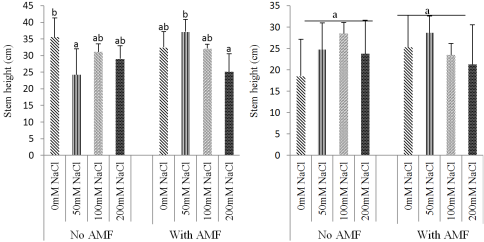
Figure 4. Effect of mycorrhizae inoculation on height of the plants under salt stress: Ricinodendron heudelotii (A) Cola acuminata (B).
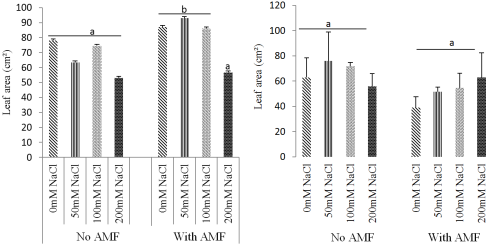
Figure 5. Effect of mycorrhizae inoculation on the leaf area as influenced by salinity: Ricinodendron heudelotii (A) and Cola acuminata (B).
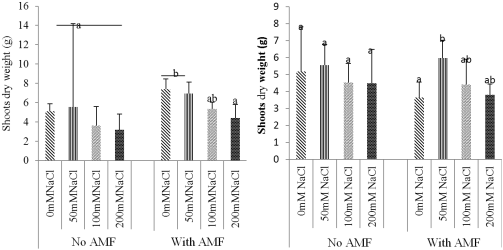
Figure 6. Effect of NaCl and biofertilizer on SDW: Ricinodendron heulotii (A) and cola acuminata (B).
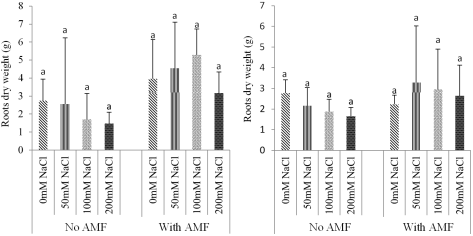
Figure 7. Effect of NaCl and biofertilizer on RDW: Ricinodendron heudelotii (A) and Cola acuminata (B).
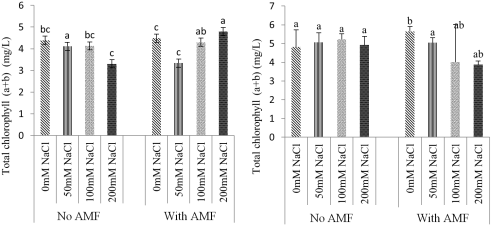
Figure 8. Effect of mycorrhizae on total chlorophyll of Ricinodendron heudelotii (A) and Cola acuminata (B) under salt stress.
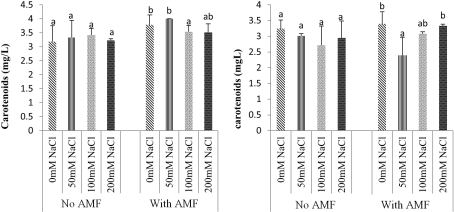
Figure 9. Effect of mycorrhizae on carotenoids content of Ricinodendron heudelotii (A) and Cola acuminata (B).
Information

